Протеорхопсин - Proteorhodopsin
Бұл мақала оқырмандардың көпшілігінің түсінуіне тым техникалық болуы мүмкін. өтінемін оны жақсартуға көмектесу дейін оны мамандар емес адамдарға түсінікті етіңіз, техникалық мәліметтерді жоймай. (Мамыр 2016) (Бұл шаблон хабарламасын қалай және қашан жою керектігін біліп алыңыз) |
| Протеорхопсин | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Протеорхопсин мультфильмінің көрнекілігі ELViture | |||||||||
| Идентификаторлар | |||||||||
| Таңба | Bac_rhodopsin | ||||||||
| Pfam | PF01036 | ||||||||
| InterPro | TCDB | ||||||||
| PROSITE | PDOC00291 | ||||||||
| SCOP2 | 2бр / Ауқымы / SUPFAM | ||||||||
| TCDB | 3.E.1 | ||||||||
| OPM суперотбасы | 6 | ||||||||
| OPM ақуызы | 4hyj | ||||||||
| |||||||||
Протеорхопсин (сонымен бірге рРодопсин) 50-ден асқан отбасы фотоактивті ретинилиден ақуыздары, үлкенірек отбасы туралы трансмембраналық ақуыздар сол пайдалану торлы қабық сияқты хромофор жарық медиациясы үшін, бұл жағдайда, а протонды сорғы. Кейбір гомологтар пентамера немесе гексамера түрінде болады.[түсіндіру қажет ] рРодопсин табылған теңіз планктоникалық бактериялар, архей және эукариоттар (прота ), бірақ бірінші болды табылды бактерияларда.[1][2][3][4]
Оның атауы шыққан протеобактериялар аталған Ежелгі грек Πρωτεύς (Протеус ), ерте теңіз құдай аталған Гомер ретінде «Теңіз қариясы «, Ῥόδος (rhódon)»Роза «, оның қызғылт түсіне байланысты және ὄψις (опсис ) «көру» үшін. Отбасының кейбір мүшелері, Гомологиялық родопсин - тәрізді пигменттер, яғни бактериорходопсин (оның 800-ден астам түрі бар) бар Сенсорлық функциялар сияқты опсиндер, үшін интеграл визуалды фототрансляция. Осы сенсорлық функциялардың көпшілігі белгісіз - мысалы, функциясы Нейропсин адамның торлы қабығында.[5] Мүшелерде әр түрлі болатыны белгілі сіңіру спектрлер, соның ішінде жасыл және көк көрінетін жарық.[6][7][8][9][10][11]
Ашу
Протеорхопсин (PR немесе рРодопсин) алғаш рет 2000 жылы а бактериялық жасанды хромосома бұрын өңделмеген теңіз γ-протеобактериялар, әлі күнге дейін тек олардың метагеномдық мәліметтер риботипі, SAR86. Зерттеулер төрт тараптың бірлескен күш-жігері болды: Oded Beja, Marcelino T. Suzuki және Эдвард Ф. Делонг кезінде Monterey Bay аквариум ғылыми-зерттеу институты (Mosslanding, CA), Л.Аравинд және Евгений V Коунин Ұлттық биотехнологиялық ақпарат орталығында (Бетезда, MD), Эндрю Хадд, Линь П. Нгуен, Стиван Б. Йованович, Кристиан М. Гейтс және Робер А Фельдман молекулярлық динамикада (Санниваль, Калифорния), ақыры Джон мен Елена Спудич микробиология және молекулалық генетика кафедрасында Хьюстондағы Техас университетінің денсаулық сақтау ғылыми орталығы. Ақуызды білдіретін Gram-протеобактериялардың грам оң және теріс түрлерінің көп екендігі анықталды.[1]
Тарату
Шығыстан протеородопсиннің экспрессиялық бактериялардың үлгілері алынды Тыңық мұхит, Орталық Солтүстік Тынық мұхит және Оңтүстік мұхит, Антарктида.[12] Кейіннен протеородопсин варианттарының гендері анықталды Жерорта теңізі, Қызыл теңіздер, Саргассо теңізі, және Жапон теңізі, және Солтүстік теңіз.[4][6]
Протеорходопсиннің нұсқалары кездейсоқ таралмайды, бірақ тереңдіктің градиенттері бойынша дисперсияны нақты сіңіру-баптауға негізделген холопротеин жүйелі; бұл негізінен сумен электромагниттік сіңіру ол толқын ұзындығын жасайды градиенттер тереңдікке қатысты. Oxyrrhis Marina Бұл Динофлагеллат жасыл сіңіргіш протеородопсинмен (L109 тобының нәтижесі) простист, олар негізінен теңіз жарығы бассейндерінде және жасыл жарық әлі де бар жағалауларда болады. Карлодиний микрум, тағы бір динолагелат, оның терең суларымен байланысты болуы мүмкін көк реттелген протеородопсинді (E109) білдіреді тік көші-қон.[3] О.Марина бастапқыда а гетеротроф дегенмен, протеородопсин функционалды түрде қатыса алады, өйткені ол ең көп бөлінген ядролық ген болды және сонымен бірге организмде біркелкі емес шашыраңқы болады, бұл кейбір органоид мембрана функциясы. Бұрын эукариоттық күн энергиясын өзгертетін жалғыз ақуыз болды I фотосистема және II фотосистема. Протеородопсиннің көптеген филаларға жол ашқан бүйірлік гендердің ауысуы әдісі деген болжам жасалды. Бактериялар, архейлер және эукариялар колонияға айналады фотикалық аймақ олар қайда жарыққа шығады; Протеорходопсин су зонасының басқа бөліктеріне емес, осы аймақ арқылы тарала алды.[3][4][9][13][14]
Таксономия
Протеорходопсин ұқсас ретинилиден ақуыздарының отбасына жатады, көбінесе оның археологиялық гомологтары галорходопсин мен бактериорхопсинге ұқсас. Родопсинді сенсорлық арқылы ашты Франц Кристиан Болл 1876 жылы.[11][15] Бактериорходопсин 1971 жылы табылып, 1973 жылы аталды және қазіргі кезде тек бактериялар емес, тек археа аймағында бар екендігі белгілі.[16] Галорходопсин алғаш рет 1977 жылы ашылып, аталды.[17] Бактериорходопсин мен галорходопсин тек Архея аймағында болады, ал протеородопсин бактерияларды, археяларды және эукариоттарды қамтиды. Протеорходопсин жеті трансмембранамен бөліседі α-спиралдар Шифф негіз механизмімен ковалентті байланысқан торлы қабық лизин жетінші спиралдағы қалдық (спираль G). Бактериорходопсин, протеородопсин сияқты, жеңіл қозғалатын протонды сорғы. Сенсорлық Родопсин - бұл көру қабілетіне қатысатын G-мен байланысқан ақуыз.[1][17]
Белсенді сайт
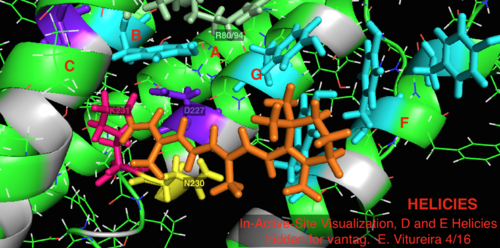
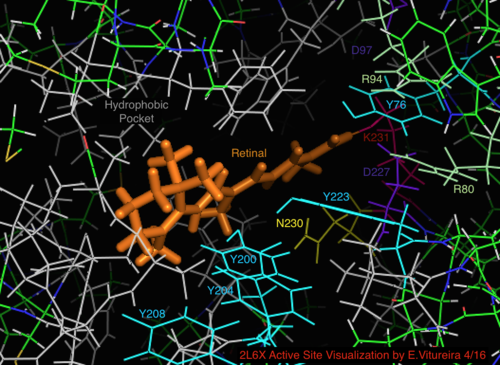
Белгілі археологиялық гомолог бактериорхопсинмен салыстырғанда, бактериородопсин механизмі үшін белгілі маңызы бар белсенді учаске қалдықтарының көп бөлігі протеородопсинде сақталады. Тізбектегі ұқсастық гало немесе бактериородопсиннен айтарлықтай сақталмайды. Гомологтары белсенді сайт қалдықтар Арг 82, Асп 85 (бастапқы протон акцептор), Асп 212 және Лис 216 ( торлы қабық Шифт базасы байланыстыратын жер) бактериорхопсинде протеородопсинде Arg94, Asp97, Asp227 және Lys231 түрінде сақталады. Алайда, протеородопсинде жоқ карбон қышқылы қалдықтары тікелей гомологты Желім Жасушадан тыс бетіндегі протонды бөлу жолына қатысады деп саналатын бактериородопсиннің 194 немесе Glu204 (немесе бакРодопсиннің нұсқасына байланысты Glu 108 және 204). Алайда, Asp97 және Arg94 бұл функцияны бактериорходопсиндегідей қалдықтың жақын орналасуынсыз алмастыра алады. Сиракуз университетінің химия кафедрасы Asp97 протон бөлетін топ бола алмайтындығын қатты көрсетті, өйткені босату аспарагин қышқылы тобы протондау жағдайында болған.[18][19][20][21]
Лиганд
Родопсин галопротеиндер тұқымдасы ретиналды лигандты бөліседі, А витамині альдегид, А витаминінің көптеген түрлерінің бірі - ретиналь біріктірілген поли-қанықпаған хромофор (полиен ) алынған жыртқыш диета немесе каротин жолымен (β-каротин 15,15'-моноксигеназа ).
Функция
Протеорходопсин бүкіл жер бетіндегі мұхиттарда бактериорхопсиндікіне ұқсас механизм арқылы жеңіл қозғалатын Н + сорғы ретінде жұмыс істейді. Бактериорходопсиндегі сияқты, торлы қабық хромофор протеородопсині ковалентті байланысқан апопротеин протонды арқылы Шифт базасы Lys231-де. Торлы хромофордың конфигурациясы фотолизденбеген протеородопсин негізінен транс-транс болып табылады[18], және изомерленеді жарықпен жарықтандырылған кезде 13-ден. Толық протеородопсин фотоциклінің бірнеше модельдері ұсынылды FTIR және ультрафиолет арқылы көрінеді спектроскопия; олар бактериорходопсиннің белгіленген циклдық модельдеріне ұқсайды.[18][20][21][22] Толық протеородопсинге негізделген фотожүйелер табылды және E. coli-де көрінді, бұл оларға торлы қабыққа немесе прекурсорларға сыртқы қажеттіліксіз ATP генерациясы үшін қосымша жарықтандырылған энергетикалық градиент қабілетін береді; PR-мен, биосинтетикалық жолмен фотопигментке арналған басқа бес ақуыздың коды.[23]
Генетикалық инженерия
Егер протеородопсинге ген енгізілген болса E. coli және торлы қабық осы түрлендірілгендерге беріледі бактериялар, содан кейін олар қосылады пигмент олардың ішіне жасуша қабығы және жарық болған кезде H + сорады. Қоңыр күлгін жарық сіңіруге байланысты айқын өзгерген колониялардың өкілі. Протондық градиенттерді мембраналық ақуыздың басқа құрылымдарын қуаттандыру үшін немесе көпіршік түріндегі органоидты қышқылдандыру үшін қолдануға болады.[1] Протеородопсин өндіретін протондық градиентті АТФ генерациясы үшін пайдалануға болатындығы одан әрі дәлелденді.[23]
Галерея

Холензим (жасыл) спиралдары А-G таңбаланған (күлгін), сонымен қатар ретиналды лиганд (қызғылт сары)

Терминалдарды көрсететін протеорхопсиннің үстіңгі көрінісі

РРодопсиннің 2L6X ақуыз құрылымының торлы қабықшамен байланысқан белсенді учаскесінің көрінісі, белсенділігі бойынша кодталған және таңбаланған түсті қалдықтар, лиганд сарғыш түсті.

2L6x Белсенді сайттағы мультфильмнің түрлі-түсті кодталған және таңбаланған көрнекілігі, D және E Helices, байқау үшін жасырылған, торлы лиганды байланыстыратын сайт




Пайдаланылған әдебиеттер
- ^ а б c г. Béjà O, Aravind L, Koonin EV, Suzuki MT, Hadd A, Нгуен Л.П., Йованович С.Б., Гейтс CM, Фельдман Р.А., Spudich JL, Spudich EN, DeLong EF (қыркүйек 2000). «Бактериялық родопсин: теңіздегі фототрофияның жаңа түрінің дәлелі». Ғылым. 289 (5486): 1902–6. Бибкод:2000Sci ... 289.1902B. дои:10.1126 / ғылым.289.5486.1902. PMID 10988064.
- ^ Lin S, Zhang H, Zhuang Y, Tran B, Gill J (қараша 2010). «Жетекшіге негізделген метатранскриптоматикалық талдаулар динофлагеллаттардағы жасырын геномдық ерекшеліктерді тануға әкеледі». Америка Құрама Штаттарының Ұлттық Ғылым Академиясының еңбектері. 107 (46): 20033–8. Бибкод:2010PNAS..10720033L. дои:10.1073 / pnas.1007246107. PMC 2993343. PMID 21041634.
- ^ а б c Slamovits CH, Okamoto N, Burri L, James ER, Keeling PJ (2011). «Теңіз эукариоттарындағы бактериялық протеородопсин протон сорғысы». Табиғат байланысы. 2 (2): 183. Бибкод:2011NatCo ... 2E.183S. дои:10.1038 / ncomms1188. PMID 21304512.
- ^ а б c Frigaard NU, Martinez A, Mincer TJ, DeLong EF (ақпан 2006). «Теңіз планктониялық бактериялары мен архейлер арасындағы протеорхопсиннің бүйірлік генінің ауысуы». Табиғат. 439 (7078): 847–50. Бибкод:2006 ж. 439..847F. дои:10.1038 / табиғат04435. PMID 16482157.
- ^ Buhr ED, Yue WW, Ren X, Jiang Z, Liao HW, Mei X, Vemaraju S, Nguyen MT, Reed RR, Lang RA, Yau KW, Van Gelder RN (20 қазан 2015). ""Нейропсин (OPN6) сүтқоректілердің торлы қабығындағы және көздің қабығындағы жергілікті циркадиандық осцилляторларды фотосуретке түсіру"". Ұлттық ғылым академиясының материалдары. 112 (42): 13093–13098. Бибкод:2015 PNAS..11213093B. дои:10.1073 / pnas.1516259112. PMC 4620855. PMID 26392540.
- ^ а б Béjà O, Spudich EN, Spudich JL, Leclerc M, DeLong EF (маусым 2001). «Мұхиттағы протеорхопсин фототрофиясы». Табиғат. 411 (6839): 786–9. дои:10.1038/35081051. PMID 11459054.
- ^ Man D, Wang W, Sabehi G, Aravind L, Post AF, Massana R, Spudich EN, Spudich JL, Béjà O (сәуір 2003). «Теңіз протеородопсиндеріндегі диверсификация және спектрлік баптау». EMBO журналы. 22 (8): 1725–31. дои:10.1093 / emboj / cdg183. PMC 154475. PMID 12682005.
- ^ Kelemen BR, Du M, Jensen RB (желтоқсан 2003). «Протеорхопсин тірі түсте: тірі бактерия жасушаларының спектрлік қасиеттерінің әртүрлілігі». Biochimica et Biofhysica Acta (BBA) - Биомембраналар. 1618 (1): 25–32. дои:10.1016 / j.bbamem.2003.10.002. PMID 14643930.
- ^ а б Sabehi G, Киркуп BC, Rozenberg M, Stambler N, Polz MF, Béjà O (мамыр 2007). «Жерорта теңізінің шығысы мен Саргассо теңізінен шығатын дивергентті теңіз протеородопсиндеріндегі бейімделу және спектрлік баптау». ISME журналы. 1 (1): 48–55. дои:10.1038 / ismej.2007.10. PMID 18043613.
- ^ Неруологиялық ғылымдар энциклопедиясы. Академиялық баспасөз. 29 сәуір 2014 ж. 441. ISBN 978-0-12-385158-1.
- ^ а б Giese, Артур С (қыркүйек 2013). Фотофизиология: жалпы қағидалар; Өсімдіктерге жарықтың әсері. Elsevier. б. 9. ISBN 978-1-4832-6227-7.
- ^ Venter JC, Remington K, Heidelberg JF, Halpern AL, Rusch D, Eisen JA, Wu D, Paulsen I, Nelson KE, Nelson W, Fouts DE, Levy S, Knap AH, Lomas MW, Nealson K, White O, Peterson J , Hoffman J, Parsons R, Baden-Tillson H, Pfannkoch C, Rogers YH, Smith HO (сәуір 2004). «Саргассо теңізінің экологиялық геномдық мылтық тізбегі». Ғылым. 304 (5667): 66–74. Бибкод:2004Sci ... 304 ... 66V. CiteSeerX 10.1.1.124.1840. дои:10.1126 / ғылым.1093857. PMID 15001713.
- ^ Джованнони, Сдж; Биббс, Л; Чо, БК; Stapels, MD; Дезидерио, Р; Вергин, KL; Раппе, MS; Лэйни, С; Вильгельм, LJ; Трипп, Х.Дж .; Матхур, Э.Дж.; Барофский, ДФ (3 қараша 2005). «SAR11 теңіз бактериясындағы протеорхопсин». Табиғат. 438 (7064): 82–5. Бибкод:2005 ж. 438 ... 82G. дои:10.1038 / табиғат04032. PMID 16267553.
- ^ Кушваха, СК; Кейтс, М (23 тамыз 1973). «Halobacterium cutirubrum ішіндегі» бактериорходопсин «мен минор С40-каротиноидтарды бөлу және идентификациялау». Biochimica et Biofhysica Acta (BBA) - липидтер және липидтер алмасуы. 316 (2): 235–43. дои:10.1016/0005-2760(73)90013-1. PMID 4741911.
- ^ Неврологиялық ғылымдар энциклопедиясы. Академиялық баспасөз. Сәуір 2014. б. 441. ISBN 978-0-12-385158-1.
- ^ Oesterhelt, D; Stoeckenius, W (29 қыркүйек 1971). «Halobacterium halobium күлгін қабығынан алынған родопсин тәрізді ақуыз». Табиғат жаңа биология. 233 (39): 149–52. дои:10.1038 / newbio233149a0. PMID 4940442.
- ^ а б Мацуно-Яги, А; Мукохата, Y (9 қыркүйек 1977). «Бактериорходопсиннің мүмкін екі рөлі; пигментациямен ерекшеленетін Halobacterium halobium штамдарын салыстырмалы зерттеу». Биохимиялық және биофизикалық зерттеулер. 78 (1): 237–43. дои:10.1016 / 0006-291x (77) 91245-1. PMID 20882.
- ^ а б c Диоумаев А.К., Браун Л.С., Ших Дж, Спудич Э.Н., Спудич Дж.Л., Лании Дж.К. (сәуір 2002). «Протеородопсиннің фотохимиялық реакция циклындағы протонды беру». Биохимия. 41 (17): 5348–58. дои:10.1021 / bi025563x. PMID 11969395.
- ^ Partha R, Krebs R, Caterino TL, Braiman MS (Маусым 2005). «Консервелген аргининнің протеородопсин хромофорымен байланысының әлсіреуі және оның қарсы әрекеті бактериорходопсиннен құрылымдық айырмашылықты білдіреді». Biochimica et Biofhysica Acta (BBA) - Биоэнергетика. 1708 (1): 6–12. дои:10.1016 / j.bbabio.2004.12.009. PMID 15949979.
- ^ а б Диумаев А.К., Ванг Дж.М., Балинт З, Варо Г, Лании Дж.К. (маусым 2003). «Протонды протеородопсинмен тасымалдау көздің торлы қабығының Schiff Asp-97 қарсы антионының анионды болуын талап етеді». Биохимия. 42 (21): 6582–7. дои:10.1021 / bi034253r. PMID 12767242.
- ^ а б Кребс Р.А., Аликиев У, Партха Р, Девита А.М., Брайман М.С. (сәуір 2002). «Протеородопсиннен жылдам жарықпен белсенді H + бөлінуі мен M аралық түзілуін анықтау». BMC физиологиясы. 2: 5. дои:10.1186/1472-6793-2-5. PMC 103662. PMID 11943070.
- ^ Xiao Y, Partha R, Krebs R, Braiman M (қаңтар 2005). «Протеородопсинмен жылдам өтпелі Н + шығаруға қатысатын фототүсірілімдердің уақыт бойынша шешілген FTIR спектроскопиясы». Физикалық химия журналы B. 109 (1): 634–41. дои:10.1021 / jp046314g. PMID 16851056.
- ^ а б Мартинес А, Брэдли А.С., Уалдбауэр JR, Summons RE, DeLong EF (2007). «Протеорхопсин фотосистемасының генінің экспрессиясы гетерологиялық иесінде фотофосфорлануға мүмкіндік береді». PNAS. 104 (13): 5590–5595. Бибкод:2007PNAS..104.5590M. дои:10.1073 / pnas.0611470104. PMC 1838496. PMID 17372221.