Қосарлы ұрықтандыру - Double fertilization


Қосарлы ұрықтандыру күрделі болып табылады ұрықтандыру гүлді өсімдіктердің механизмі (ангиоспермдер ). Бұл процесс әйелдің қосылуын қамтиды гаметофит (мегагаметофит, деп те аталады эмбрион қап ) екі еркекпен гаметалар (ұрық). Бұл а тозаң дәні стигмасын ұстанады карпель, гүлдің аналық көбею құрылымы. Тозаң дәні содан кейін ылғалды қабылдап бастайды өніп шығады, қалыптастыру тозаң түтігі бағытына қарай созылады аналық без стиль арқылы. Содан кейін тозаң түтігінің ұшы аналық безге еніп, арқылы енеді микропил ұрықта ашылу. Тозаң түтігі мегагаметофиттегі екі сперматозоидты бөліп шығарады.
Ұрықтанбаған жұмыртқа жасушаларының саны 8-ден тұрады және 3 + 2 + 3 түрінде орналасқан (жоғарыдан төмен), яғни 3 антиподальды жасушалар, 2 полярлық орталық жасушалар, 2 синергидтер және 1 жұмыртқа жасушалары. Бір сперматозоидтар жұмыртқа клеткасын ұрықтандырады, ал екінші сперматозоидтар екеуімен қосылады полярлық ядролар үлкендер орталық ұяшық мегагаметофиттің Гаплоидты сперматозоидтар мен гаплоидты жұмыртқа қосылып а түзеді диплоидты процесс деп аталады сингамия, ал басқа сперматозоидтар және екеуі гаплоидты мегамаметофиттің үлкен орталық жасушасының полярлық ядролары а түзеді триплоидты ядро (үш есе біріктіру). Кейбір өсімдіктер пайда болуы мүмкін полиплоид ядролар. Гаметофиттің үлкен жасушасы кейін дамиды эндосперм, дамушы эмбрионды тамақтандыратын қоректік заттарға бай тін. Аналық без, ұрықшаларды қоршап, ұрықтарды қорғайтын жемістерге айналады және оларды тарату үшін жұмыс істеуі мүмкін.[1]
Эндоспермаға ықпал ететін екі орталық жасушалық аналық ядролар (полярлық ядролар) митоз арқылы пайда болады. мейоздық жұмыртқаны тудырған өнім. Триплоидты эндоспермнің генетикалық конституциясына аналық үлес эмбрионның қосқан үлесімен тең.
Зауытта 2008 жылы жүргізілген зерттеуде Arabidopsis thaliana, аналық жыныс жасушаларында еркек ядроларының миграциясы, аналық ядролармен бірігу кезінде алғаш рет қолданылып құжатталды in vivo бейнелеу. Миграция мен синтез процесіне қатысатын кейбір гендер де анықталды.[2]
Жылы қосарланған ұрықтандырудың дәлелі Гнеталес, олар гүлденбейтін тұқым өсімдіктері болып саналады.[3]
Қысқа тарих
Қосарлы ұрықтандыруды бір ғасырдан астам уақыт бұрын ашқан Сергей Навашчин және Григнард Киев,[4] Ресей империясы, және Леон Гиньяр Франция. Әрқайсысы бір-біріне тәуелсіз жаңалық ашты.[5] Лилиум мартагоны және Фритиллария тенелла классикалық әдіспен жасалған қосарланған ұрықтандырудың алғашқы бақылауларында қолданылды жарық микроскопы. Жарық микроскопының шектеулеріне байланысты қосарлы ұрықтандыру процесіне қатысты көптеген жауапсыз сұрақтар туындады. Алайда, дамуымен электронды микроскоп, көптеген сұрақтарға жауаптар берілді. Ең бастысы, В.Дженсен тобы жүргізген бақылаулар еркек жыныс жасушаларында жоқ екенін көрсетті жасуша қабырғалары және бұл плазмалық мембрана гаметалар тозаң дәнінің ішінде оларды қоршап тұрған жасушаның плазмалық мембранасына жақын.[6]
Гимноспермадағы қосарланған ұрықтандыру
Қосарланған ұрықтандырудың әлдеқайда рудиментарлы түрі гнеталес ретінде белгілі гимноспермдер қатарының жыныстық көбеюінде орын алады.[3] Нақтырақ айтсақ, бұл оқиға екеуінде де құжатталған Эфедра және Гнетум, ішкі бөлігі Гнетофиттер.[7] Жылы Ephedra nevadensis, бір бинокулалық сперматозоидтар жұмыртқа жасушасына түседі. Бастапқы ұрықтану оқиғасынан кейін екінші сперматозоид ядросы жұмыртқа цитоплазмасында табылған қосымша жұмыртқа ядросын ұрықтандыру үшін бағытталады. Көптеген басқа тұқымдық өсімдіктерде бұл екінші «вентральды каналдың ядросы» функционалдық тұрғыдан жарамсыз болып табылады.[8] Жылы Gnetum гнемоны, көптеген бос жұмыртқа ядролары әйелдің ішіндегі цитоплазмасында бар гаметофит. Жетілген аналық гаметофиттің енуінен сәттілік тозаң түтігі, аналық цитоплазма және бос ядролар тозаң түтігін қоршап қозғалады. Бинуклеаттық сперматозоидтан босатылған екі сперматозоидтар, содан кейін бос жұмыртқа ядроларымен қосылып, өміршең екі зиготалар түзеді, бұл отбасылар арасындағы гомологиялық сипаттама. Эфедра және Гнетум.[9] Екі отбасында да ұрықтандырудың екінші оқиғасы қосымша диплоид түзеді эмбрион. Бұл супер сандық эмбрион кейіннен үзіліп, тек бір жетілген эмбрионның синтезіне әкеледі.[10] Қосымша ұрықтандыру өнімі Эфедра алғашқы эмбрионды тамақтандырмайды, өйткені аналық гаметофит қоректік заттармен қамтамасыздандыруға жауап береді.[9] Қосарланған ұрықтандырудың қарабайыр процесі гимноспермалар нәтижесінде бір жұмыртқа жасушасында орналасқан екі диплоидты ядро пайда болады. Бұл ерекшеленеді ангиосперма жағдай, бұл жұмыртқа жасушасының бөлінуіне әкеледі және эндосперм.[11] Геномындағы салыстырмалы молекулалық зерттеулер Gnemon гнетофиттердің неғұрлым тығыз байланысты екенін анықтады қылқан жапырақты ағаштар олар ангиоспермаларға қарағанда.[12][13][14] Гнеталиялар мен ангиоспермдерді апалы-сіңлілі таксондар деп анықтайтын антофиттік гипотезадан бас тарту қосарланған ұрықтану процесінің өнімі деген болжамға әкеледі. конвергентті эволюция және гнетофиттер мен ангиоспермалардың арасында дербес пайда болды.[15]
Экстракорпоральды ұрықтандыру
Экстракорпоральды ұрықтандыру көбінесе молекулалық өзара әрекеттесуді, сондай-ақ гүлді өсімдіктердегі гаметалар синтезінің басқа аспектілерін зерттеу үшін қолданылады. Еркек пен аналық жыныс жасушалары арасындағы экстракорпоральды ұрықтандыруды дамытудағы негізгі кедергілердің бірі - сперматозоидтардың тозаң түтігінде және эмбрион қапшығында жұмыртқаның ұсталуы. Жұмыртқа мен сперматозоидтардың бақыланатын біріктірілуіне қазірдің өзінде қол жеткізілді көкнәр өсімдіктері.[16] Тозаңның өнуі, тозаң түтігінің енуі және екі рет ұрықтандыру процестері қалыпты түрде жүретіні байқалды. Іс жүзінде бұл әдіс әртүрлі гүлді өсімдіктерде тұқым алу үшін қолданылған және оны «пробирка арқылы ұрықтандыру» деп атаған.[17]
Байланысты құрылымдар мен функциялар
Мегагаметофит
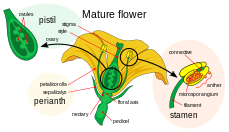
Қос ұрықтандыруға қатысатын аналық гаметофит - мегагаметофит ангиоспермдер гаплоидты, эмбрион қапшығы деп аталады. Бұл an ішінде дамиды жұмыртқа, а негізінде аналық без жатыр карпель. Мегагаметофиттің айналасы (бір немесе) екі бүтіндер, деп аталатын саңылау құрайды микропил. Мегагаметофит, әдетте гаплоидты, (-дан) басталады диплоидты ) мегаспора аналық жасуша, деп те аталады мегаспороцит. Келесі оқиғалар тізбегі белгілі бір түрге байланысты өзгеріп отырады, бірақ көптеген түрлерде келесі оқиғалар болады. Мегаспороцит төрт гаплоидты мегаспора түзіп, мейоздық жасуша бөлінуіне ұшырайды. Нәтижесінде пайда болған төрт мегаспораның біреуі ғана тірі қалады. Бұл мегаспора митоздық бөлінудің үш кезеңінен өтеді, нәтижесінде сегіз гаплоидты ядролары бар жеті жасуша пайда болады (орталық жасушада полярлық ядролар деп аталатын екі ядро бар). Эмбрион қапшығының төменгі ұшы басқа екі гаплоидты жасушаның ортасында орналасқан гаплоидты жұмыртқа жасушасынан тұрады. синергидтер. Синергидтер тозаң түтігін мегропаметофитке тарту және микропиле арқылы бағыттауда қызмет етеді. Мегагаметофиттің жоғарғы ұшында антиподальды жасушалар орналасқан.
Микрогаметофит
Қос ұрықтандыруға қатысатын аталық гаметофиттер немесе микрогаметофиттер құрамында болады тозаң астық. Олар микроспорангия шеңберінде дамиды, немесе тозаң қаптары стаменттердегі тозаңдардың Әрбір микроспорангиумда диплоид бар микроспора аналық жасушалар немесе микроспороциттер. Әрбір микроспороцит мейозға түсіп, төрт гаплоидты микроспора түзеді, олардың әрқайсысы соңында тозаң дәніне айналуы мүмкін. Микроспора өтеді митоз және цитокинез екі бөлек жасушаны, яғни генеративті және түтікшелі жасушаны шығару үшін. Бұл екі жасуша спора қабырғасынан басқа жетілмеген тозаң дәнін құрайды. Еркек гаметофит жетіле келе генеративті жасуша түтікшелі жасушаға өтіп, генеративті жасуша митозға түсіп, екі ұрық жасушасын түзеді. Тозаң дәні пісіп болғаннан кейін тозаңқаптар тозаңды босатып, ашыңыз. Тозаңды дейін жеткізеді пистилла жел немесе жануарлардың тозаңдандырғышымен және стигмаға түскен басқа гүлден. Тозаң дәні өнген сайын, түтік жасушасы созылып, карпельдің ұзын стилінде созылып, аналық безге дейін созылатын тозаң түтігін шығарады, онда оның сперматозоидтары мегагаметофитте шығарылады. Қосарлы ұрықтандыру осы жерден басталады.[18]
Сондай-ақ қараңыз
Әдебиеттер тізімі
- ^ Бергер, Ф. (қаңтар 2008). «Қос ұрықтандыру, мифтерден шындыққа дейін». Өсімдіктің жыныстық көбеюі. 21 (1): 3–5. дои:10.1007 / s00497-007-0066-4. S2CID 8928640.
- ^ Бергер, Ф .; Хамамура, Ю. & Ингуф, М. және Хигашияма, Т. (тамыз 2008). «Қосарлы ұрықтандыру - заңмен ұсталды». Өсімдіктертану тенденциялары. 13 (8): 437–443. дои:10.1016 / j.tplants.2008.05.011. PMID 18650119.CS1 maint: бірнеше есімдер: авторлар тізімі (сілтеме)
- ^ а б В.Рагхаван (қыркүйек 2003). «Қосарлы ұрықтандыру туралы, оның ашылуынан қазіргі уақытқа дейінгі кейбір ойлар». Жаңа фитолог. 159 (3): 565–583. дои:10.1046 / j.1469-8137.2003.00846.x.
- ^ Kordium EL (2008). «[Гүлді өсімдіктерде қосарлы ұрықтану: 1898-2008]». Цитол. Генет. (орыс тілінде). 42 (3): 12–26. PMID 18822860.
- ^ Дженсен, В.А. (ақпан 1998). «Қосарлы ұрықтандыру: жеке көрініс». Өсімдіктің жыныстық көбеюі. 11 (1): 1–5. дои:10.1007 / s004970050113. S2CID 33416360.
- ^ Dumas, C. & Rogowsky, P. (тамыз 2008). «Ұрықтану және тұқымның ерте қалыптасуы». Comptes Rendus Biologies. 331 (10): 715–725. дои:10.1016 / j.crvi.2008.07.013. PMID 18926485.
- ^ Кармайкл, Дж. С .; Фридман, В.Э. (1995-12-01). «Gnetum гнемонындағы қосарлы ұрықтандыру: жасуша циклі мен жыныстық көбею арасындағы байланыс». Өсімдік жасушасы. 7 (12): 1975–1988. дои:10.1105 / tpc.7.12.1975. ISSN 1040-4651. PMC 161055. PMID 12242365.
- ^ Фридман, Уильям Э. (1990). «Ephedra nevadensis (Ephedraceae) кезіндегі жыныстық көбею: гүлденбейтін тұқым өсімдігінде қосарлы ұрықтанудың қосымша дәлелі». Американдық ботаника журналы. 77 (12): 1582–1598. дои:10.1002 / j.1537-2197.1990.tb11399.x. JSTOR 2444491.
- ^ а б Кармайкл, Джеффри С .; Фридман, Уильям Э. (1996). «Gnetum гнемонындағы (Gnetaceae) қосарлы ұрықтандыру: оның гнеталес пен антофит кладындағы жыныстық көбею эволюциясы». Американдық ботаника журналы. 83 (6): 767–780. дои:10.1002 / j.1537-2197.1996.tb12766.x. JSTOR 2445854.
- ^ Фридман, В.Э. (1995-04-25). «Органикалық қайталану, фитнес теориясы және альтруизм: эндоспермнің эволюциясын және ангиоспермдік репродуктивті синдромды түсіну». Ұлттық ғылым академиясының материалдары. 92 (9): 3913–3917. Бибкод:1995 PNAS ... 92.3913F. дои:10.1073 / pnas.92.9.3913. ISSN 0027-8424. PMC 42072. PMID 11607532.
- ^ Фридман, Уильям Э. (1994). «Тұқымдық өсімдіктердегі эмбриогенияның эволюциясы және эндоспермнің шығу тегі мен ерте тарихы». Американдық ботаника журналы. 81 (11): 1468–1486. дои:10.1002 / j.1537-2197.1994.tb15633.x. JSTOR 2445320.
- ^ Боу, Л.Мишель; Пальто, Гвинея; деПамфилис, Клод В. (2000-04-11). «Барлық үш геномдық бөліктерге негізделген тұқым өсімдіктерінің филогенезі: экстенсивті гимноспермалар монофилетикалық, ал Гнеталестің ең жақын туыстары - қылқан жапырақты ағаштар». Ұлттық ғылым академиясының материалдары. 97 (8): 4092–4097. Бибкод:2000PNAS ... 97.4092B. дои:10.1073 / pnas.97.8.4092. ISSN 0027-8424. PMC 18159. PMID 10760278.
- ^ Қыс, Кай-Уве; Беккер, Аннет; Мюнстер, Томас; Ким, Ян Т .; Саедлер, Хайнц; Тиссен, Гюнтер (1999-06-22). «MADS-box гендері гнетофиттердің гүлді өсімдіктерге қарағанда қылқан жапырақты ағаштармен тығыз байланысты екенін анықтайды». Ұлттық ғылым академиясының материалдары. 96 (13): 7342–7347. Бибкод:1999 PNAS ... 96.7342W. дои:10.1073 / pnas.96.13.7342. ISSN 0027-8424. PMC 22087. PMID 10377416.
- ^ Магаллон, С .; Сандерсон, Дж. (2002-12-01). «Жоғары консервіленген гендерден алынған тұқымдық өсімдіктер арасындағы қатынастар: ежелгі тұқымдар арасында қарама-қайшы филогенетикалық сигналдарды сұрыптау». Американдық ботаника журналы. 89 (12): 1991–2006. дои:10.3732 / ajb.89.12.1991 ж. ISSN 1537-2197. PMID 21665628.
- ^ Чау, Шу-Миав; Паркинсон, Кристофер Л .; Чэн, Ючан; Винсент, Томас М .; Палмер, Джеффри Д. (2000-04-11). «Үш өсімдік геномынан алынған тұқым өсімдіктерінің филогениясы: сақталған гимноспермалардың монофилиясы және қылқан жапырақты ағаштардан Гнеталестің шығу тегі». Ұлттық ғылым академиясының материалдары. 97 (8): 4086–4091. Бибкод:2000PNAS ... 97.4086C. дои:10.1073 / pnas.97.8.4086. ISSN 0027-8424. PMC 18157. PMID 10760277.
- ^ Зенктелер, М. (1990). «Жоғары сатыдағы өсімдіктерде экстракорпоральды ұрықтандыру және кең будандастыру». Crit Rev Plant Sci. 9 (3): 267–279. дои:10.1080/07352689009382290.
- ^ Рагхаван, В. (2005). Қосарлы ұрықтандыру: гүлді өсімдіктердегі эмбрион мен эндоспермнің дамуы (суретті ред.). Бирхязер. 17-19 бет. ISBN 978-3-540-27791-0.
- ^ Кэмпбелл Н.А.; Reece JB (2005). Биология (7 басылым). Сан-Франциско, Калифорния: Pearson Education, Inc., 774–777 бет. ISBN 978-0-8053-7171-0.
| Пәндер | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Зауыт топтар | |||||||||||
| |||||||||||
| |||||||||||
| Өсімдіктің өсуі және әдеті | |||||||||||
| Көбейту | |||||||||||
| Өсімдіктер таксономиясы | |||||||||||
| Тәжірибе | |||||||||||
| |||||||||||
| |||||||||||