Поликрикос - Polykrikos
| Поликрикос | |
|---|---|
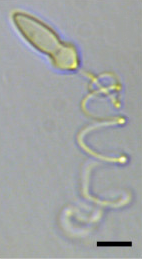 | |
| A жарық микрограф туралы Polykrikos kofoidii экструдты көрсету нематоциста. Масштаб жолағы = 10µм.[1] | |
| Ғылыми классификация | |
| (ішілмеген): | |
| (ішілмеген): | |
| Филум: | |
| Сынып: | |
| Тапсырыс: | |
| Отбасы: | |
| Тұқым: | Поликрикос |
Поликрикос (грек тілінен аударғанда «поли» - көп, ал «крикос» - сақина немесе шеңбер) - отбасы тектілерінің бірі Поликрикас құрамына атекатты псевдоколония түзуші динофлагеллаттар кіреді. Поликрикос күрделі баллистикалық аппаратпен сипатталады,[2] түрлердің әртүрлі организмдерге жем болуына мүмкіндік беретін нематоцист-таениоцист кешені деп аталды. Поликрикос балдырлардың гүлденуін реттейтіні анықталды, өйткені олар улы динофлагеллаттармен қоректенеді.[3][4] Сонымен қатар, Поликрикос туралы балыққа улы болатын кейбір мәліметтер бар.[5]
Білім тарихы
Поликрикос алғаш рет 1868 жылы Ульянин көрді және қателесіп турбелярлық жалпақ құрттардың метазоан личинкасы ретінде қарастырылды. 1873 жылы Бутшили үлгіні қайта зерттеп, жасуша ерекше кирпик болды деген қорытындыға келді, ал Берг кейінірек, 1881 ж. Поликрикос динофлагеллаттарға жақындықтар.[6]
Морфология
Поликрикос біртұтас жасушаның бір уақытта қызмет атқаратын зоооидтар колониясы (отарлық организмнің бірліктері).
Барлық Поликрикос түрлерге ие: 1) ағзаның артқы ұшына дейін созылған сәл қисық бойлық борозда, сулькус 2) цикл тәрізді акробаза, бұл сулькадан алдыңғы кеңею болып табылады 3) көлденең бороз, цингул, ығысумен 4) таениоцист -нематоциста кешендері 5) зоооидтерге қарағанда ядролардың санынан екі-төрт есе аз, және 6) зоооидтары аз және бір ғана ядросы бар псевдоколонияларға бөлшектеу қабілеті.[7]
Бұл тұқымның айрықша ерекшелігі - зооидтардың жұп санынан тұратын көп ядролы псевдоколониялардың түзілуі. Әрбір зооидта жұп флагелла бар (көлденең және бойлық флагелла) және өзінің көлденең ойығы, цингуласы бар, бірақ зооидты бойлық бороздар, сульци біріктірілген.[8] Көлденең флагеллада басқа динофлагеллаттарға ортақ бүйірлік проекциялар, мастигоналар және жолақты жіп бар.[9] Жиі Поликрикос зоооидтерге қарағанда ядролардың жарты санына ие, және зоооидтардың әр жұбы ядроға ие. Топ ішінде органоидтар көрсетілген бірнеше вариация бар, бірақ таксон ішінде әр түрлі мүшелерден трихоцисталар, нематоцисталар, таениоциттер, мукоцисталар және пластидтер байқалды.[8]
Цитоплазмасы Поликрикос көптеген өрескел эндоплазмалық торлармен, Гольджи кешендерімен және вакуолаларымен сипатталады. Поликрикос экструзомалар шығаратын органоидтар шығаратыны белгілі. Олардың бірі - зооидтарда түзілген нематоциста. Ағзада кездесетін тағы бір экструзом - таяқша тәрізді таениоциста, ол нематоцистаға дистальды орналасқан және бұрын қателесіп нематоцист-прекурсор ретінде қарастырылған.[10] Бұл органеллалар бірігіп, ең жақсы синапоморфия деп саналатын таениоцист-нематоциста кешенін құрайды. Поликрикос қаптау.[8] Гольджиден алынған вакуольдер екі органеллалармен бөліседі және әрқайсысына NTC артикуляциясына қатысумен бірге оның өсуіне қажет молекулалар береді.[10] Органеллалар жақын жерде орналасқан, бірақ әр түрлі мембраналарда орналасқан және «шұңқыр» деп аталатын үзіндімен бөлінген. Нематоциста - үлкен органелл және таениоцистаның артында жатыр. Кейбір соңғы зерттеулер [3] екі органеллалардың жұмысы біріктіріліп, таениоциста жемге жабысады, содан кейін нематоцист разряды жыртқыштың тесілуіне әкеледі және ақырында артқы весикулаға жақын нематоцисталардың соңында орналасқан сүйрейтін жіптің көмегімен олжаны алады. Нематоцистаға енген түтік жемге қарай ағып кетеді және жемді тесуге арналған деп жорамалдайды.
Сонымен қатар, Гавелис және басқалар.[3] терең зерттелген NTC морфологиясы және баллистикалық механизм, олар книдарийлерден түбегейлі ерекшеленді, нематоцисталар бір клеткалы динофлагеллаттарда дербес дамыды. Киндрийлерден айырмашылығы, қапталған түтікшелі капсула мөрмен жабылған, бұл стилетті ату кезінде алдымен жіптен босату үшін капсуланы ішінен тесуге, кейінірек жыртқышты тесуге мәжбүр етеді. Түтікше саптамадан өтіп бара жатқанда, ол оперулуланы ашады және ширатылады. Книдарийлердегі нематоцисталардағы баллистика осмостық қоздырғыш поли гамма глутамат синтазасы PgsAA синтезімен қозғалады, ал Поликрикос бұл капсула қабырғасындағы капсула талшығының жиырылуы нәтижесінде туындаған қысым деп саналады.[3]Ядро екі қабатты талшықты қабықпен ерекшеленеді, ол эвинацияланған ядролық қабықтың астын сызады; кортекс ядроға күш пен форма беру үшін гипотеза жасайды, ал ядролық эвагинациялар кортекстің тесілу орындарында ядролық-цитоплазмалық алмасу аймағын ұлғайтады деп есептеледі.[9]
Жылы Поликрикос, дәл анықталған талшықты ленталар ядролық-флагеллярлық байланыстарға қатысады, ал флагеллярлық аппаратураға бекіту қозғалыс, митоз және клеткалардың бөліну процестері кезінде ядро флагеллаларға қатысты бағытталуы мүмкін.[9]
Пластидтер
Арасында қоректік заттарды алудың вариациясы бар Поликрикос түрлер, өйткені кейбіреулері тек фотосинтезге сүйенеді, ал кейбіреулері - микотрофтар, ал кейбіреулері - міндетті гетеротрофтар. Поликрикос органеллярлық эволюцияны зерттеуге арналған пайдалы топ. Ерте тармақталған поликрикоидтар, Polykrikos geminatum және P. hartmanii, үш қабатты қабаттасқан тилакоидтары бар үш қабатты пластидтер бар, олар динофлагеллаттар үшін жалпы қайталама перидинин типті пластидтерді көрсетеді. Алайда, микотрофты P. lebouriae, гетеротропты поликрикоидтар арасында филогенетикалық ұя салған, динофлагеллаттардың типтік емес пластидтері бар. Оның екі қабығы бар және құрамында диатомалар мен гаптофиттерде кездесетін екі қабатты тилакоидтар бар. Алайда, молекулалық деректерді Гавелис және басқалармен талдау.[11] мұны көрсетті P. lebouriae перидинин типті пластидтерге ие, олар, бәлкім, ата-баба поликрикоидтарынан алынған. Транскриптомиктерді талдау поликрикоидтардағы көптеген шығындар оқиғаларын көрсетті, оларды энергетика және физиологиялық шектеулер тұрғысынан түсіндіруге болады. Мөлшерін біртіндеп ұлғайту Поликрикос организмдерді керемет жыртқыш етеді, бірақ бір жасушалардың дене мөлшерінің ұлғаюы хлоропластардың өздігінен көлеңкеленуіне әкеліп соғады және фотосинтездеу тиімділігінің төмендеуіне алып келеді. Мамандандырылған NTC және жасушалардың үлкен мөлшері фотосинтездің бірнеше рет жоғалуына себеп болуы мүмкін.[11]
Өмірлік циклі P. kofoidii
Үшін Поликрикос, түрдің көбеюі туралы толық мәліметтер қол жетімді (голотип) P. kofoidii, оның өмірлік циклі жалпы динофлагеллаттар цикліне ұқсайды, өйткені вегетативті жасушалар цистол жасай алатын диплоидты (2n) зигота түзетін гаметалар түзеді, бірақ псевдоколониялық табиғат бірқатар ерекшеліктер қосады Поликрикос даму.[12]
Организмдер жақсы тамақтанғанда, олар 4-зоид-2-ядролардың псевдоколониялары түрінде пайда болды, ал вегетативті көбею кезінде зоооидтар саны екі еселеніп, содан кейін ядролар бөлініп, 8-зооид-4-ядролар сатысына алып келеді, әрі қарай көлденең екілік екіге бөлінеді. зооид-2-ядролар Поликрикос. Гаметаның түзілуі ерекше болды, өйткені псевдоколония вегетативті жасушаларға қарағанда әртүрлі мөлшердегі және морфологиялы 4 гаметаларды түзеді. Вегетативті форма екі еселенген зооидтарды құрады және кейіннен 2-зооид-1-ядро түріндегі төрт гаметаға бөлінеді. Екі гаметалар вентральды бүйірлерімен одан әрі жұптасып, планозиготаны біріктірді. Үшін P. kofoidii гаметаларда түйіспелі саусақ тәрізді екі құрылым байқалды, олар гаметалармен жанасуға және бірігуге қатысады, бірақ мұны растау үшін көбірек мәліметтер қажет. Вентральды түрде біріктірілген гаметалар сегіз флагелланың күрделі қайта құрылуын және сульци мен цингулидің түзілуін қажет етті. 4-зооидты планозиготаның бір ғана ядросы болды және тағамның қол жетімділігіне байланысты екі даму жолы болды. Планозигота аштық жағдайында бір ядросы жетіспейтін екі 2-зооидқа бөлшектелген, бірақ одан әрі тағдыр зерттелген жоқ. Мәдениет жағдайында көптеген организмдер мейозға ұшырап, вегетативті циклге тікелей енген. Киста сатысында өте аз планозиготалар өтті. Киста кезеңі 1 ай бойы сақталды, бұл тыныштық кезеңі 6 айға дейін жетуі мүмкін басқа динофлагеллаттармен салыстырғанда салыстырмалы түрде қысқа мерзім деп саналады. Циклдың ұзақтығы экологиялық қоректену стратегиясымен байланысты. Қысқа тыныштық кезеңі азық-түліктің қол жетімділігі өзгерген жағдайда гетеротрофты түрлерге пайдалы өмірлік цикл кезеңдерінің арасындағы жылдам велосипед айналымын жеңілдетуі мүмкін. Алайда, кистаны шығаруға қатысатын химиялық сигнал беру мүмкіндігі Поликрикос әлі анықталуы керек.
Гермлинг, цистадан шыққан бір зоидті жасуша, ешқашан еркін тіршілік ететін динофлагеллаттар үшін ешқашан жазылмаған ерекше дамуға ие болды. Оның морфологиясы 1-зооид-1-ядродан, 2-зооид-1-ядродан және 4-зооид-1-ядродан 4-зооид-2-ядро сатысына өтті. Бұл деректер осындай штрихтау әдісі көрініс бере ала ма деген сұрақ тудырады Поликрикос псевдоколониялар филогениясы.[12]
Тіршілік ортасы және экология (тауашасы, тамақ)
Барлығы белгілі Поликрикос түрлері теңіз орталарында кездеседі. Түрлердің көпшілігі планктонды, себебі оларды алып тастайды Polykrikos lebouriae және Polykrikos herdmanae бентикалық тіршілік ету ортасында кездеседі.[7]Сондай-ақ, тамақтану экологиясының өзгеруі бар, өйткені кейбір түрлерде пластидтер бар және олар қоректік заттар алу үшін фотосинтезді қолдана алады, бірақ көбінесе миксотрофтар болып келеді (P. barnegatensis, P. lebouriae, P tanit, P. hartmannii). Тек бір түрдің тек автотрофты екендігі белгілі (P. geminatum), ал кейбіреулері пластидтен тұрады және толығымен гетеротрофты (P. grassei, P. herdmanae, P. kofoidii және P. Schwartzii),.[13][8]
Polykrikos hartmannii фототрофты динофлагеллат болып табылады және Канада суларында белгілі болды,[14] АҚШ, Мексика, Қытай, Үндістан, Жапония, Корея ,.[5][13] P. hartmanii нематоцист-таениоцист кешенін (кейінірек НТК деп атайды) пайдаланып, жемтігіне якорь салғаннан кейін балдыр түрлерімен қоректенеді.[13] Тан және басқалар.[5] байқалды P. hartmannii кәмелетке толмаған қой қойларынан 100% өлім тудырған гүлдену (Ципринодон) 24 сағат ішінде P. hartmannii ихтиотоксикалық, зиянды балдыр болып табылады. P.hartmannii тізбек түзетін динофлагеллаттармен қоректенеді Кохлодиний поликрикоидтары және Gymnodinium catenatum олар сонымен бірге балықтардың өлімін тудыратыны белгілі, сондықтан P. hartmannii осы жыртқыш динофлагеллаттар өндіретін токсиндерді залалсыздандыратын ферменттері бар деп саналады.[13] Сондай-ақ, үшеуін салыстыру Поликрикос түрлердің қоректенуі түрлердің өздеріне деген сүйіспеншілігімен ерекшеленетінін, ал кейбіреулері басқаларына қарағанда анағұрлым мамандандырылған екенін көрсетті P. hartmanii жыртқыштарға қарағанда азырақ әр түрлі (2 жыртқыш түрмен қоректенеді) P. kofoidii және P. lebouriae14 түрлі балдыр түрлерімен қоректенетін.[13]
Гетеротрофты жыртқыштық Поликрикос организмдердің кейбіреулері улы гүлденуді тудыратын динофлагеллаттарда жайылып жүргендіктен үлкен қызығушылық тудырды. Жыртқыштықтың жоғары әсері Polykrikos schwartzii Ботшили улы динофлагеллатта Александрия (Lebour) Balech туралы Аргентинада хабарланды, ал Polykrikos kofoidii Чаттон басқарды Гимнодий катенатумы Грэм Португалия мен Жапонияның жағалау суларында.[4] G. catenatum - паралитикалық ұлулармен улануды тудыратын түрлердің бірі (PSP) және Австралия, Жапония, Мексика және Испания суларында кездеседі.[15] Тамақтану тәртібін бақылаулар ұсынады P. kofoidii бастапқыда оның ілмегіне жақын циклмен жүзу тәртібі байқалды, содан кейін нематоциста ағып, артқы сулькус арқылы жыртқыштың денеге тартылуы және жыртқыштың соңғы жұтылуы.[16]PSP әлеуметтік-экономикалық-экологиялық мәселелерді көтереді, өйткені бұл теңіз сүтқоректілерінің де, адамдардың да денсаулығына, сондай-ақ улы микробалдырлардың популяциясын реттеу механизміне әсер етеді. P.kofoidii қоршаған ортаны бақылау мен денсаулыққа қауіпті жоюда маңызды болуы мүмкін.
Тәжірибелік маңыздылығы
Гетеротрофты динофлагеллаттармен улы микробалдырлардың жыртылуы - балдырлардың гүлденуін бақылайтын факторлардың бірі. Поликрикос сияқты динофлагеллаттар популяциясын модуляциялайтыны белгілі Александрия,[3][4] және G. catenatum,[17] олар улы балдырлардың гүлденуінің кең таралған агенттері болып табылады. Мұндай гетеротрофты поликрикоидтар теңіздегі қоректік торларда олардың жемтігі тудыратын уыттылық деңгейін төмендетіп қана қоймай, улы гүлденуді тоқтатып, биоремедиацияда қолдануға болады.[17] Осылайша, судың уыттылығының төмендеуі теңіздегі қоректік торлардың тепе-теңдігін реттеуге және балықтардың, теңіз сүтқоректілерінің және теңіз құстарының өлім-жітімін төмендетуге көмектеседі. Детоксикацияның молекулалық механизмдерін одан әрі зерттеу Поликрикос биомедициналық және қоршаған ортаны бақылау саласында пайдалы болуы мүмкін.
Алайда, кейбіреулер Поликрикос белгілі бір балықтарға денсаулыққа қауіп төндіреді, ал гүлденуді реттейтіндер көбіне теңіз омыртқасыздары сияқты жыртқыш болады. амфиподтар токсиндерді қайтадан тамақтану желісіне қайтарады.[17] Кейбір поликрикоидтар популяциясы мониторингі және бақташылардың организміндегі токсиндер динамикасын зерттеу планктонға негізделген тамақ торларын жақсы түсінуге, экожүйелердегі улану дәрежесін бағалауға және токсиндерді жоюдың әлеуетті жолдарын ұсынуға мүмкіндік береді.
Түрлер тізімі
Қазіргі уақытта келесі 10 түр таксономиялық жолмен қабылданады: Polykrikos barnegatensis Мартин Г.В. Polykrikos geminatus (F.Schütt) D.X.Qiu & Senjie Lin, Polykrikos grassei Лекальды, Polykrikos hartmannii В.Зиммерманн,Polykrikos herdmaniae Хоппенрат және Леандр,Polykrikos kofoidii Чаттон,Polykrikos lebouriae Гердман, Polykrikos schwartzii Бютчли, Polykrikos tanit Рене,Polykrikos tentaculatus О.Ветцель [18]
Әдебиеттер тізімі
- ^ Хоппенрат, Мона; Бахваров, Цветан Р; Ыңғайлы, Сара М; Делвич, Чарльз Ф; Leander, Brian S (2009). «SSU және LSU рДНК тізбектерінен алынған, құрамында окселлоидты динофлагеллаттардың (Warnowiaceae) молекулалық филогениясы». BMC эволюциялық биологиясы. 9: 116. дои:10.1186/1471-2148-9-116. PMC 2694157. PMID 19467154.
- ^ Джеймс Горман (31 наурыз 2017). «Іс-әрекеттегі бір жасушалы арпунердің алғашқы айқын көрінісі». NYTimes.com. Алынған 26 желтоқсан 2017.
- ^ а б c г. e Гавелис, Г .; Уакеман, К .; Рипкен, С .; Озебек, С .; Гольштейн, Т .; Херранц М .; Килинг, П .; Leander, B. (2017). «Микробтық қару жарысы: динофлагеллаттардағы баллистикалық» нематоцисталар «органеллалардың күрделілігінде жаңа экстремалды білдіреді». Ғылым жетістіктері. 3 (3): e1602552. дои:10.1126 / sciadv.1602552. PMC 5375639. PMID 28435864.
- ^ а б c Нагай, С .; Мацуяма, Ю .; Такаяма, Х .; Kotani, Y. (2002). «Морфология Polykrikos kofoidii және P. Schwartzii (Dinophyceae, Polykrikaceae) кисталар мәдениетте алынған ». Фикология. 41 (4): 319. дои:10.2216 / i0031-8884-41-4-319.1.
- ^ а б c Тан, Ю.З .; Харке, МДж .; Gobler, CJ (2013). «Морфологиясы, филогениясы, динамикасы және ихтиотоксичность Pheopolykrikos hartmannii (Dinophyceae) оқшаулайды және Нью-Йорктен гүлдейді, АҚШ ». Дж.Фикол. 49 (6): 1084–1094. дои:10.1111 / jpy.12114.
- ^ Тейлор, Ф.Ж.Р (1987). «Динофлагеллаттар биологиясы». Биологиялық монографиялар. 21.
- ^ а б Хоппенрат, М .; Леандр, Б.С. (2007б). «Псевдоколониялық динофлагеллаттардың морфологиясы және филогениясы Polykrikos lebourae және Polykrikos herdmanae n. sp «. Протист. 158: 209–227. дои:10.1016 / j.protis.2006.12.001.
- ^ а б c г. Хоппенрат, М .; Leander, B. S. (2007a). «Поликрикоидты динофлагеллаттардағы сипаттамалық эволюция». Дж.Фикол. 43: 366–377. дои:10.1111 / j.1529-8817.2007.00319.x.
- ^ а б c Брэдбери, П.К .; Вестфолл, Дж .; Таунсенд, Дж. (1983). «Динофлагелланың ультрақұрылымы Поликрикос. Ядроның және оның флагман аппаратурасымен байланысы ». Ультрақұрылымдық зерттеулер журналы. 85 (1): 24–32. дои:10.1016 / s0022-5320 (83) 90113-2. PMID 6686618.
- ^ а б Вестфолл, Дж. А .; Брэдбери, П.С .; Таунсенд, Дж. (1983). «Динофлагелланың ультрақұрылымы Поликрикос. Нематоцист-таениоциста кешенін және экструзияға арналған орынның морфологиясын дамыту ». J ұялы ғылыми жұмыс. 63: 245–261.
- ^ а б Гавелис, Г .; Уайт, Р.А .; Саттл, C. А .; Килинг, П.Ж .; Leander, B. S. (2015). «Біріктірілген ПЦР көшбасшысын қолданатын бір клеткалы транскриптомика: поликрикоидты динофлагеллаттардағы фотосинтездің бірнеше рет жоғалуының дәлелі». BMC Genomics. 16 (1). дои:10.1186 / s12864-015-1636-8. PMC 4504456. PMID 26183220.
- ^ а б Тиллманн, У .; Хоппенрат, М. (2013). «Псевдоколониялық динофлагелланың өмірлік циклі Polykrikos kofoidii (Gymnodiniales, Dinoflagellata) ». Фикология журналы. 49 (2): 298. дои:10.1111 / jpy.12037. PMID 27008517.
- ^ а б c г. e Ли, Дж .; Джонг, Х. Дж .; Ли, К.Х .; Джанг, С. Х .; Ким Дж. Х .; Ким, К.Ю. (2015). «Нематоциста-таэниокиста құрамындағы фототрофты динофлагеллааттағы микотрофия Polykrikos hartmannii". Зиянды балдырлар. 49: 124–132. дои:10.1016 / j.hal.2015.08.006.
- ^ Хоппенрат, М .; Юбуки, Н .; Бахваров, С .; Leander, B. S. (2010). «Қайта жіктеу Pheopolykrikos hartmannii сияқты Поликрикос (Dinophyceae) ішінара күрделі экструзомдардың ультрақұрылымына негізделген ». Еуропа. Дж. Протистол. 46: 29–37. дои:10.1016 / j.ejop.2009.08.003. PMID 19767184.
- ^ Мацуяма, Ю; Миямото, М; Kotani, Y (1999). «Гетеротрофты динофлагеллаттың жайылымдық әсері Polykrikos kofoidii гүлдену кезінде Gymnodinium catenatum". Aquat Microb Ecol. 17: 91–98. дои:10.3354 / ame017091.
- ^ Мацуока, К .; Чо, Х. Дж .; Джейкобсон, Д.М. (2000). «Гетеротрофты динофлагеллаттардың тамақтану тәртібін және өсу қарқынын бақылау, Polykrikos kofoidii (Polykrikaceae. Dinophyceae) 39: 82- 86 ». Фикология. 39: 82–86. дои:10.2216 / i0031-8884-39-1-82.1.
- ^ а б c Чжон Х .; Парк, К .; Ким Дж .; Канг, Х .; Ким, С .; Чой, Х., ...; Парк, М. (2003). «Динофлагелланның уыттылығының төмендеуі Gymnodinium catenatum гетеротрофты динофлагеллатпен қоректенгенде Polykrikos kofoidii". Су микробтарының экологиясы. 31: 307–312. дои:10.3354 / ame031307.
- ^ Гури, М. Д. «Поликрикос Бутчли, 1873». Балдырлар. Алынған 20 ақпан, 2017.