Лепидодендрон - Lepidodendron
| Лепидодендрон | |
|---|---|
 | |
| The стробилус туралы Лепидодендрон | |
| Ғылыми классификация | |
| Корольдігі: | Планта |
| Клайд: | Трахеофиттер |
| Клайд: | Ликофиттер |
| Сынып: | Ликоподиопсида |
| Тапсырыс: | †Лепидодендралдар |
| Отбасы: | †Лепидодендрея |
| Тұқым: | †Лепидодендрон Штернберг |
| Түрлер | |
| |
Лепидодендрон болып табылады жойылған түр қарабайыр, тамырлы, ағаш тәрізді өсімдіктер, деп те аталады ағаштар, байланысты квиллворт және ликопсидтер (клубтық мүктер). Олар көмір орманы флора. Олар кейде 50 метр биіктікке (160 фут) жетті,[1] магистральдар көбінесе диаметрі 1 метрден (3,3 фут) асатын. Олар кезінде өркендеді Көміртекті Кезең (шамамен 359,2 ± 2,5 Мя (миллион жыл бұрын дейін табылды Кейінгі триас, шамамен 205 Mya) жойылмас бұрын.[2] Кейде қате түрде «алып клуб мүктері» деп аталатын, бұл тұқым заманауи клубтық мүктерге қарағанда қазіргі заманғы квиллворлармен тығыз байланысты болды.
Этимология
Аты Лепидодендрон шыққан Грек λεπίς лепис, масштаб және δένδρον дендрон, ағаш.
Сипаттамасы және биологиясы

Лепидодендрон түрлері қазіргі заманғы ағаштармен салыстыруға болатын. Өсімдіктің негізінде ені 2 м (6,6 фут) болатын конустық діңдер болған, олар шамамен 40 м (130 фут) дейін көтерілген.[3] және тіпті 50 м (160 фут),[1] көптеген тамырлармен жабылған көлденең тарайтын бұтақтардың жерасты жүйесінен туындайды. Ағаштардың биіктігі өсімдіктерді қазіргі ағаштарға ұқсас етсе де, тұрақты дихотомия филиалдар құрылды әдет бұл қазіргі заманғы ағаштарға қайшы келеді. Бұтақтардың ұштарында сопақ тәрізді болды конустар а-ның қазіргі конустарына ұқсас пішінді болды шырша немесе шырша.[4]
Ағаштардың сабағында бет пішіні жоқ болатын тамырлы камбий, қазіргі заманғы ағаштардың қос тамырлы камбийінен айырмашылығы. Қазіргі заманғы ағаштардың қос беттік камбиясы екіншісін де береді флоэма және ксилема, бір беткейлік камбий Лепидодендрон ағаштар тек екінші ксилема шығарды. Ағаштар қартайған кезде бір беткейлік камбиум өндіретін ағаш өсімдіктің жоғарғы жағына қарай төмендеді, сондықтан түпкі бұтақтар жасқа ұқсайды Лепидодендрон сабақтар. Ағаштардың сабақтарында және бұтақтарында қазіргі заманғы ағаштармен салыстырғанда аз ағаш болды, ал жетілген сабақтарының көп бөлігі массивтен тұрады кортикальды меристема. Бұл кортикальды тіннің біркелкі өсуі жыл мезгілдерінің өзгеруінде және ұйқының болмауында ешқандай айырмашылық жоқ екенін көрсетеді бүршіктер бұдан әрі жоқтығын көрсетеді маусымдық жылы Лепидодендрон түрлері.[5] Ескі сабақтардың сыртқы қабығы қабықшаға айналған ликоподиопсид перидерма.[6] Ағаштардың қабығы біршама ұқсас болды Пицея түрлері, сияқты жапырақ шрамы қабық созылған сайын созылып, жыртылып тұратын қазық тәрізді проекциялар қалыптасты. Желдің иілу күшіне қарсы тұру үшін, Лепидодендрон ағаштар тамырлардың ұлпаларына емес, сыртқы қабығына тәуелді болды, бұл қазіргі заманғы ағаштармен салыстырғанда, көбінесе олардың орталық массасына сүйенеді.[4]
Ағаштардың жапырақтары ине тәрізді болды және олардың әрқайсысында жалғыз дана болатын өсінділерге тығыз спираль тәрізді болды тамыр. Жапырақтары кейбір түрлеріндегі шыршаның жапырақтарына, ал жапырақтарына ұқсас болды Pinus roxburghii басқаларында, дегенмен жалпы Лепидодендрон түрлерін олардың түрлерінен ажыратуға болмайды Сигиллария түрлері. The анық емес жапырақтары бұтақтардың айналасында цилиндрлік қабықша құрады. Жапырақтары тек жіңішке және жас бұтақтарда болды, бұл ағаштар әрқашан жасыл болғанымен, олар өздерінің инелерін қазіргі қылқан жапырақты ағаштар сияқты ұзақ уақыт сақтамайтындығын көрсетті. Жапырақ жастықшалары фюзиформалы және ұзартылған, ұзындығы 8 см (3,1 дюйм) және ені 2 см (0,79 дюйм) дейін өскен. Жапырақ жастықтарының ортасы тегіс болды, қайда жапырақ шрамы болған кезде жасалған абстиссия жапырақ оның түбінен кесіп тастайды. Әрбір жапырақ шрамы орталық дөңгелек немесе үшбұрышты тыртықтан және кішірек және сопақ тәрізді екі бүйірлік тыртықтан тұрды. Бұл орталық шрам негізгі қай жерде екенін көрсетеді тамырлы шоқ тамырдың тамыр жүйесіне байланысты жапырақтың. Бұл ксилема байламы тек бастапқыдан құралған трахея. Екі сыртқы шрамдар тамырлы тіннің тармағынан өтіп кеткен шанышқы тармақтарын белгілейді қыртыс жапырақтағы сабақтың. Бұл айыр бұрымды кейде «парихнос» деп те атайды. Бұл жіптің айналасында болды паренхима жасушалар және кейде қалың қабырғалы элементтер. Екі өткізгіш тіннің айналасында трансфузияның кең қабығы болды трахеидтер. Жапырақ тыртықшасының астында базальды жағдайға қарай жастықша жастықшасы жұмсарған. Бұл тарылған аймақта ұңғыма шұңқырлары бар дөңгелек әсерлер болды. Бұл әсер тарылтылған бөліктің жоғарғы жағындағы парихнос тыртықтарымен үздіксіз болды. Бұл әсерді қалыптастырады аэренхима парихноспен тығыз дамыған тін. Жапырақ шрамының үстінде тереңдік үшбұрышты әсер «ұқсастығы шұңқыр» ретінде белгілі болды лигула туралы Isoetes. Кейбір жапырақты жастықшаларда буын шұңқырының үстінде екінші депрессия болды. Оның мақсаты түсініксіз болғанымен, депрессия а позициясын белгілеуі мүмкін деген болжам жасалды спорангиум. А Лепидодендрон Ағаш жапырақ жастығы белгілі мөлшерде өскен, жапырақ жастығы созылған. Бұл созылу жапырақты жастықшаларды бөліп тұрған ойықты кеңейтіп, кең, тегіс канал жасады.[4]
Гифалар кейде тіндерде болады Лепидодендрон бейімділікті көрсететін ағаштар саңырауқұлақ паразиттер.[4]
Әр түрлі қазба ыдыраудың әр түрлі деңгейлерін атауға сипатталған Лепидодендрон қабығының сүйектері. Аты Бержерия эпидермистен айрылған сабақтарды сипаттайды, Аспидиариу жастықтар терең ыдырап жойылған кезде қолданылады, және Кноррия жапырақ жастықшалары мен кортикальды тіндердің көп бөлігі шіріп кеткен кезде қолданылады, ал таяз «флют» беті қалады. Алайда, бұлар қабықтың сақталған түрлеріне қарағанда өсу формалары болуы мүмкін деген болжам жасалды, өйткені біркелкі пішіндермен бүтіндей қазылған діңдер табылды; егер ыдырау магистраль бойында тұрақты деп есептелсе, онда әртүрлі формалар ыдырау деңгейіне емес, өсуге нұсқайды. Мүмкін, оның магистралі Лепидодендрон ағаштар өсу формаларына ұшырады Кноррия, Аспидиария, және Бержерия сәйкесінше магистральды жоғарылату.[7]
Көбейту
Лепидодендрон түрлердің өсу циклінен тұратын 10-15 жыл өмір сүру циклі болған, онда ағаштар алдын-ала белгіленген биіктікке дейін өскен және одан кейінгі репродуктивті цикл, онда ағаштар репродуктивті мүшелер шығарды, содан кейін ағаштар өмірге ұқсас өлді а циклі Mauna Kea күміс сөз.[6]
Тұқыммен көбейтудің орнына, Лепидодендрон споралармен көбейетін ағаштар. Споралар сақталды спорангиялар негізгі магистральда немесе оның жанында өсетін құнарлы сабақтарда орналасқан. Құнарлы сабақтар бұтақтардың ұштарында шоғырланған конус тәрізді құрылымдарда бірге өскен.[8]
Тарату
Жетіспеушілігі өсу сақиналары және ұйықтап жатқан бүршіктер маусымдық өсу заңдылықтарын көрсетпейді және ұқсас сипаттамалары бар заманауи өсімдіктер өсуге бейім тропикалық шарттар, бірақ Лепидодендрон түрлері бүкіл аумаққа таратылды субтропикалық шарттар. Ағаштар сол уақыттағы тропикалық флорамен салыстырғанда кең аумақты мекендеді, ағаштар солтүстікке қарай өскен Шпицберген және оңтүстікке қарай Оңтүстік Америка, ішінде ендік диапазоны 120 °.[5]
Төмендеу және жойылу
Бойынша Мезозой дәуірде алып ликопсидтер жойылып, олардың орнына қылқан жапырақты ағаштар да, кішілері де пайда болды квиллворт.[2]Бұл жаңа пайда болған ағаштың бәсекелестігінің нәтижесі болуы мүмкін гимноспермалар. Лепидодендрон - кең таралған өсімдіктердің бірі қазба қалдықтары табылды Пенсильвания -жас (Кейінгі көміртек ) жыныстар. Олар басқа жойылған ликопсидтер тұқымдастарымен тығыз байланысты, Сигиллария және Лепидендропсис.
Бұл бөлім кеңейтуді қажет етеді. Сіз көмектесе аласыз оған қосу. (Мамыр 2012) |
Бұқаралық мәдениетте
19 ғасырда гауһар тәрізді жапырақты тыртық өрнегінің рептилия көрінісіне байланысты, тасқа айналған магистральдар Лепидодендрон әуесқойлар жәрмеңке алаңдарында алып қазба ретінде жиі қойылды кесірткелер немесе жыландар.[9] Керісінше, а темноспондил Тасманиядағы триас формациясында жиналған қосмекенділер 1885 жылы ежелгі өсімдіктің конусы ретінде сипатталған, осы түрге жақындықтармен бірге Lepidostrobus muelleri.[10][11]
Галерея

Сыртқы пішіні Лепидодендрон бастап Жоғарғы көміртекті туралы Огайо
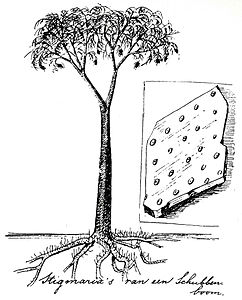
Қайта құру Лепидодендрон голландиялық табиғатты қорғаушы Эли Хейманс (1911)

Лепидодендрон дисплейде Пенсильвания штатының мемлекеттік мұражайы



Сондай-ақ қараңыз
Әдебиеттер тізімі
- ^ а б В. В. Алехин (1961). Geografiia rastenii s osnovani botaniki (Өсімдіктер географиясы және ботаника негіздері). Господин педагог-педагог. изд-во. б. 167. Алынған 2020-10-05.
- ^ а б Sahney, S., Benton, MJ & Falcon-Lang, HJ (2010). «Тропикалық орманның құлауы Пенсильваниядағы тетраподты әртараптандыруға түрткі болды». Геология. 38 (12): 1079–1082. дои:10.1130 / G31182.1.CS1 maint: бірнеше есімдер: авторлар тізімі (сілтеме)
- ^ Лопатин А. (2012). Палеонтологический музей имени Ю.А. Орлова (Орлов палеонтология мұражайы). Мәскеу: PIN RAN. б. 56. ISBN 978-5-903825-14-1. Алынған 2020-10-05.
- ^ а б c г. Сьюард, Альберт Чарльз (1898). Қазба өсімдіктері: ботаника және геология студенттеріне арналған. 1. Кембридж университетінің баспасы. 93–192 бет.
- ^ а б Вульф, Евгений Владимирович және Бриссенден, Элизабет (1943). Тарихи өсімдіктер географиясына кіріспе. Chronica Botanica компаниясы. 176–177 бб.CS1 maint: бірнеше есімдер: авторлар тізімі (сілтеме)
- ^ а б Карл Дж. Никлас (1997). Өсімдіктердің эволюциялық биологиясы (суретті ред.). Чикаго Университеті. б. 321. ISBN 9780226580838.
- ^ Томас, Б.А. және Уотсон, Джоан (1976). «Болтоннан, Ланкаширден алынған 114 футтық лепидодендрон». Геологиялық журнал. Wiley онлайн кітапханасы. 11 (1): 15–20. дои:10.1002 / gj.3350110102.CS1 maint: бірнеше есімдер: авторлар тізімі (сілтеме)
- ^ Джон Адам Дорр, Дональд Ф. Эшман (1970). Мичиган геологиясы (суретті ред.). Мичиган университеті. б. 429. ISBN 9780472082803.
- ^ Роберт Логан Джек (1886). Квинсленд геологиясының анықтамалығы. Квинслендтің геологиялық қызметі. Уорвик пен Сапсфорд. б. 28.
- ^ Джонстон, Р.М. (1885). «Кампанияның құмтастарында конустың, лепидостробтардың бір түрінің табылуы». Тасмания корольдік қоғамының құжаттары мен еңбектері. Қоғам. 1884: 225.
- ^ Розефельдс, А.С.; Уоррен, А. (Қыркүйек 2011). "Lepidostrobus muelleri Джонстон - бұл темноспондилді қосмекенділердің бас сүйегінің үзіндісі ». Альчеринга: Австралия палеонтология журналы. 35 (3): 459–462. дои:10.1080/03115518.2011.527163. S2CID 83845584.
Әрі қарай оқу
- Дэвис, Пол; Kenrick, Paul (2004). Қазба өсімдіктері. Вашингтон, ДС: Смитсондық кітаптар. ISBN 1-58834-181-X.
- Морран, Робин С. (2004). Папоротниктердің табиғи тарихы. Портланд: Timber Press. ISBN 0-88192-667-1.
- Кристофер Дж. Клил мен Барри А. Томастың «Британдық көмір шаралары бойынша өсімдік қалдықтары», пабл. Палеонтологиялық қауымдастық, Лондон, 1994 ж., 222 бет, ISBN 0-901702-53-6
- Дж.М.Андерсон және Х.М.Андерсон. 1985. Оңтүстік Африканың палеофлорасы. Оңтүстік Африка Мегафлорасының Девонионнан төменгі борға дейінгі продромы 1-423