Иммуноглобулиннің ауыр тізбегі - Википедия - Immunoglobulin heavy chain
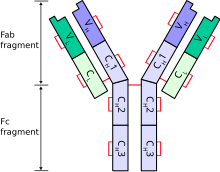

The иммуноглобулиннің ауыр тізбегі (IgH) - бұл үлкен полипептидті суббірлік антидене (иммуноглобулин). Адам геномында IgH генінің локустары 14 хромосомада орналасқан.
Әдеттегі антидене екі иммуноглобулиннен (Ig) және екі тізбектен тұрады Ig жеңіл тізбектері. Ауыр тізбектің бірнеше түрі бар, олар класты немесе изотип антидене. Бұл ауыр тізбектің түрлері әр түрлі жануарларда әр түрлі болады. Барлық ауыр тізбектер қатарынан тұрады иммуноглобулин домендері, әдетте бір айнымалысы бар домен (VH) байланыстыру үшін маңызды антиген және бірнеше тұрақты домендер (CH1, CH2 және т.б.). Тіршілікке қабілетті ауыр тізбекті өндіру В клеткасының жетілуіндегі басты қадам болып табылады. Егер ауыр тізбек суррогат жеңіл тізбегімен байланысып, плазмалық мембранаға ауыса алса, онда дамып келе жатқан В клеткасы өзінің жеңіл тізбегін жасай бастайды.[2]
Ауыр тізбек әрқашан жеңіл тізбекті байланыстыра бермейді. В-ге дейінгі лимфоциттер ауыр тізбекті жеңіл тізбек болмаған кезде синтездей алады, содан кейін ауыр тізбекті ауыр тізбекті байланыстыратын ақуызбен байланыстыруға мүмкіндік береді.[3]
Сүтқоректілерде
Сабақтар
Сүтқоректілердің иммуноглобулиннің ауыр тізбегінің бес түрі бар: γ, δ, α, μ және ε.[4] Олар анықтайды сыныптар иммуноглобулиндер: IgG, IgD, IgA, IgM және IgE сәйкесінше.
- Ауыр тізбектерде α және 450 шамамен 450 аминқышқылдары бар.
- Ауыр тізбектерде μ және 550 шамамен 550 аминқышқылдары бар.[4]
Аймақтар
Әрбір ауыр тізбектің екі аймағы бар:
- а тұрақты аймақ (бұл бір кластағы барлық иммуноглобулиндер үшін бірдей, бірақ кластар арасында әр түрлі).
- а айнымалы аймақ арасында ерекшеленеді В жасушалары, бірақ бірдей В жасушасы немесе В жасушасы шығаратын барлық иммуноглобулиндер үшін бірдей клон. Кез-келген ауыр тізбектің айнымалы саласы бір иммуноглобулиндік доменнен тұрады. Бұл домендердің ұзындығы шамамен 110 амин қышқылдары.[6]
Сиырлар
Сиырлар, арнайы Бос таурус, ауыр тізбек болатын жалпы сүтқоректілер тақырыбының вариациясын көрсетіңіз CDR H3 аймағы антиденелердің дивергентті репертуарын шығаруға бейімделген, олар таныс валентті ұшы бетінің орнына антигеннің өзара әрекеттесу бетін «сабақ пен тұтқаны» ұсынады.[7] Сиыр CDR-сі ерекше ұзын және жұптастырылған өндірісті қолдайтын бірізділік атрибуттарын қамтиды цистеин кезінде қалдықтар соматикалық гипермутация.[7] Осылайша, адамдарда соматикалық гипермутация қадамы мақсатты болады V (D) J рекомбинациясы процесс, сиырлардағы мақсат алуан түрлі болып табылады дисульфидті байланыстар және антигенмен өзара әрекеттесетін бірегей ілмектер жиынтығының генерациясы.[7] Бұл вариацияның болжамды эволюциялық драйвері - сиырдың ас қорыту жүйесінде олардың болуы нәтижесінде әр түрлі микробтық ортаның болуы. күйіс қайыратын малдар.[7]
Балықта
Жақау балықтар сүтқоректілер үшін сипатталғандай антиденелер жасауға қабілетті ең алғашқы жануарлар болып көрінеді.[8] Алайда, балықтарда сүтқоректілерде болатын антиденелердің бірдей репертуары жоқ.[9] Қазірге дейін Ig ауыр үш тізбегі анықталды сүйекті балық.
- Бірінші анықталған μ (немесе) болды му) барлық иекті балықтарда болатын және алғашқы иммуноглобулин деп санайтын ауыр тізбек. Алынған антидене, IgM, а түрінде бөлінеді тетрамер жылы телеост әдеттегідей емес, балық бесбұрыш сүтқоректілер мен акулаларда кездеседі.[дәйексөз қажет ]
- IgD үшін ауыр тізбек (δ) бастапқыда анықталды канал сомы және Атлантикалық лосось және қазір көптеген телеосттар үшін жақсы құжатталған.[10]
- Үшінші телеост Ig ауыр тізбегінің гені жақында анықталды және сүтқоректілер үшін осы уақытқа дейін сипатталған ауыр тізбектердің біріне ұқсамайды. Бұл екеуінде де анықталған ауыр тізбек радуга форелі (τ)[11] және зебрбиш (ζ),[12] мүмкін антидене түзуі мүмкін изотип (IgT немесе IgZ), олар эволюциялық тұрғыдан IgM-дан бұрын болуы мүмкін.
Сүйекті балықтар үшін байқалған жағдайға ұқсас үш ауыр Ig тізбекті изотиптері анықталды шеміршекті балықтар. Μ қоспағанда, Ig ауыр тізбекті изотиптері шеміршекті балықтарға ғана тән көрінеді. Алынған антиденелер IgW (IgX немесе IgNARC деп те аталады) және IgNAR (иммуноглобулин жаңа антиген рецепторы).[13][14] Соңғы түрі - а ауыр тізбекті антидене, жеңіл тізбектері жоқ антидене және оны өндіру үшін қолдануға болады бір доменді антиденелер, олар мәні бойынша өзгермелі домен болып табылады (VNAR) IgNAR.[15][16][17] Акуланың бір доменді антиденелері (VNARs) ісікке немесе вирустық антигендерге үлкен аңғал медбике акуладан V оқшаулауға боладыNAR кітапхананы пайдалану фагтарды көрсету технологиясы.[16]
IgW сонымен қатар целакант пен өкпе балықтарын қоса алғанда, лоб тәрізді балықтар тобында табылған. Целаканттағы IgW1 және IgW2 әдеттегі (VD) n-Jn-C құрылымына ие, сонымен қатар көптеген тұрақты домендерге ие.[18][19]
Қосмекенділерде
Бақалар IgX пен IgY синтездей алады. [20]
Сондай-ақ қараңыз
Әдебиеттер тізімі
- ^ «Мұрағатталған көшірме». Архивтелген түпнұсқа 2007 жылы 19 сәуірде. Алынған 20 сәуір, 2007.CS1 maint: тақырып ретінде мұрағатталған көшірме (сілтеме)[толық дәйексөз қажет ]
- ^ Märtensson, I-L; Ceredig, R (2017-01-23). «Суррогаттық жарық тізбегінің және В-жасуша алдындағы рецептордың тышқанның В-жасушасының дамуындағы рөлі». Иммунология. 101 (4): 435–441. дои:10.1046 / j.1365-2567.2000.00151.x. ISSN 0019-2805. PMC 2327112. PMID 11122446.
- ^ Хаас, Ингрид Г .; Вабль, Матиас (1983). «Иммуноглобулиннің ауыр тізбекті байланыстыратын ақуызы». Табиғат. 306 (5941): 387–9. Бибкод:1983 ж.т.306..387H. дои:10.1038 / 306387a0. PMID 6417546.
- ^ а б в Janeway Калифорния, кіші.; т.б. (2001). Иммунобиология (5-ші басылым). Garland Publishing. ISBN 0-8153-3642-X. (NCBI Bookshelf арқылы электронды толық мәтін).[бет қажет ]
- ^ Вуф, Дженни М .; Бертон, Деннис Р. (2004). «Адам антиденесі - кристалдық құрылымдармен жарықтандырылған Fc рецепторларының өзара әрекеттесуі». Табиғатқа шолу Иммунология. 4 (2): 89–99. дои:10.1038 / nri1266. PMID 15040582.
- ^ «Биология жобасы». Антидене құрылымы. Аризона университеті. Алынған 27 мамыр, 2020.
- ^ а б в г. Ван, Фэн; Экиерт, Дамиан С .; Ахмад, Инша; Ю, Вэнли; Чжан, Ён; Базирған, Омар; Торкамани, Әли; Раудсепп, Терье; Мванги, Вайтаса; Крисичиелло, Майкл Ф .; Уилсон, Ян А .; Шульц, Питер Г. Smider, Vaughn V. (2013). «Антиденелердің әртүрлілігін өзгерту». Ұяшық. 153 (6): 1379–93. дои:10.1016 / j.cell.2013.04.049. PMC 4007204. PMID 23746848.
- ^ Балықтың ауыр тізбегі мен жеңіл тізбекті гендері[толық дәйексөз қажет ] Мұрағатталды 23 наурыз 2007 ж Wayback Machine
- ^ Бенгтен, Ева; Клем, Л. Уильям; Миллер, Норман В .; Уарр, Григорий В.; Уилсон, Мелани (2006). «Арналық сом иммуноглобулиндері: репертуар және экспрессия». Дамытушылық және салыстырмалы иммунология. 30 (1–2): 77–92. дои:10.1016 / j.dci.2005.06.016. PMID 16153707.
- ^ Солем, Стайн Торе; Стенвик, Йорген (2006). «Телеосттарда антидене репертуарының дамуы - лосось және Гадус морхуа L-ға баса назар аудару». Дамытушылық және салыстырмалы иммунология. 30 (1–2): 57–76. дои:10.1016 / j.dci.2005.06.007. PMID 16084588.
- ^ Хансен, Дж. Д .; Ландис, Э.Д .; Филлипс, Р.Б. (2005). «Радуга фореліндегі бірегей Ig ауыр тізбекті изотиптің (IgT) ашылуы: телеост балықтарында В клеткасының ерекше даму жолының салдары». Ұлттық ғылым академиясының материалдары. 102 (19): 6919–6924. Бибкод:2005PNAS..102.6919H. дои:10.1073 / pnas.0500027102. JSTOR 3375456. PMC 1100771. PMID 15863615.
- ^ Данилова, Надия; Бусман, Джерен; Джекош, Керстин; Штайнер, Лиза А (2005). «Зеброфишадағы иммуноглобулиннің ауыр тізбекті локусы: Бұрын белгісіз изотиптің, Z иммуноглобулинінің идентификациясы және экспрессиясы». Табиғат иммунологиясы. 6 (3): 295–302. дои:10.1038 / ni1166. PMID 15685175.
- ^ Дули, Х .; Флажник, М.Ф. (2006). «Шеміршекті балықтардағы антидене репертуарының дамуы». Дамытушылық және салыстырмалы иммунология. 30 (1–2): 43–56. дои:10.1016 / j.dci.2005.06.022. PMID 16146649.
- ^ Симмонс, Дэвид П .; Абрегу, Фиона А .; Кришнан, Уша V .; Пролл, Дэвид Ф .; Стрельцов, Виктор А .; Жұмыртқа, Лариса; Хаттарки, Меган К .; Nuttall, Stewart D. (2006). «IgNAR акуласының біртекті домендік антидене фрагменттерінің өлшемдерінің стратегиялары». Иммунологиялық әдістер журналы. 315 (1–2): 171–84. дои:10.1016 / j.jim.2006.07.019. PMID 16962608.
- ^ Весоловский, Януш; Алзогарай, Ванина; Рейельт, қаңтар; Унгер, Мэнди; Хуарес, Карла; Уррутия, Мариела; Кауэрф, Ана; Данкуа, Уэлбек; Риссиек, Бьорн; Шеуплейн, Феликс; Шварц, Николь; Адриух, Сахил; Бойер, Оливье; Семан, Мишель; Лицея, Алексей; Серрезе, Дэвид V .; Голдбаум, Фернандо А .; Хааг, Фридрих; Кох-Нольте, Фридрих (2009). «Бірыңғай домендік антиденелер: инфекция мен иммунитеттегі тәжірибелік және терапевтік құралдар». Медициналық микробиология және иммунология. 198 (3): 157–74. дои:10.1007 / s00430-009-0116-7. PMC 2714450. PMID 19529959.
- ^ а б Фэн, Минцян; Биан, Хеджяо; Ву, Сяолин; Фу, Тяньюн; Фу, Ин; Хонг, Джессика; Флеминг, Брайан Д .; Флажник, Мартин Ф .; Хо, Митчелл (2019 қаңтар). «Алғашқы фен-акулалар акулаларынан алынған бір фазалы антиденелер кітапханасының үлкен фаг-бейнеленген VNAR кітапханасын құру және келесі буын тізбегін талдау». Антидене терапиясы. 2 (1): 1–11. дои:10.1093 / abt / tby011. ISSN 2516-4236. PMC 6312525. PMID 30627698.
- ^ Ағылшын, Хеджяо; Хонг, Джессика; Хо, Митчелл (2020). «Ежелгі түрлер заманауи терапевтік әдістер ұсынады: акула VNAR біртекті домендік антиденелер тізбегін, фаг кітапханаларын және ықтимал клиникалық қосымшаларын жаңарту». Антидене терапиясы. 3: 1–9. дои:10.1093 / abt / tbaa001.
- ^ Чжан, Тяньи; Такки, Лука; Вэй, Джигуо; Чжао, Яофенг; Салинас, Айрин (2014). «Африка өкпе балықтарындағы иммуноглобулиннің ауыр тізбекті гендерінің классішілік әртараптандырылуы». Иммуногенетика. 66 (5): 335–51. дои:10.1007 / s00251-014-0769-2. PMC 4348116. PMID 24676685.
- ^ Ота, Т .; Раст, Дж. П .; Литман, Г.В .; Амемия, C. Т. (2003). «Дипной ішіндегі қарабайыр иммуноглобулиннің ауыр тізбекті изотипінің тегі бойынша шектелуі эволюциялық парадоксты анықтайды». Ұлттық ғылым академиясының материалдары. 100 (5): 2501–6. Бибкод:2003PNAS..100.2501O. дои:10.1073 / pnas.0538029100. PMC 151370. PMID 12606718.
- ^ Ду, Кристина С .; Масоф, Сара М .; Крисситиелло, Майкл Ф. (2012). «Африкалық тырнақты бақаның ауызша иммунизациясы (Xenopus laevis) шырышты қабықша иммуноглобулинін IgX реттейді». Ветеринариялық иммунология және иммунопатология. 145 (1–2): 493–8. дои:10.1016 / j.vetimm.2011.10.019. PMC 3273591. PMID 22100190.
{kind=link}
{kind=link}
Сыртқы сілтемелер
- Иммуноглобулин + ауыр + тізбектер АҚШ ұлттық медицина кітапханасында Медициналық тақырып айдарлары (MeSH)
- Ауыр тізбекті талдауға арналған білім беру ресурсы