Дилофозавр - Dilophosaurus
| Дилофозавр | |
|---|---|
 | |
| Қайта құрылған актерлер құрамы голотип үлгісі (UCMP 37302) жерлеу жағдайында, Онтарионың Корольдік мұражайы | |
| Ғылыми классификация | |
| Корольдігі: | Анималия |
| Филум: | Chordata |
| Клайд: | Динозаврия |
| Клайд: | Сауришия |
| Клайд: | Теропода |
| Отбасы: | †Dilophosauridae |
| Тұқым: | †Дилофозавр Уэллс, 1970 |
| Түрлер: | †D. ветерилли |
| Биномдық атау | |
| †Dilophosaurus wetherilli (Уэллс, 1954) | |
| Синонимдер | |
| |
Дилофозавр (/г.aɪˌлoʊfəˈс.rəс,-foʊ-/[1] dy-LOHF-о-СОР-әс ) Бұл түр туралы теропод динозаврлар қазіргі өмір сүрген Солтүстік Америка кезінде Ерте юра, шамамен 193 миллион жыл бұрын. Ішінде үш қаңқа табылды солтүстік Аризона 1940 ж., ал ең жақсы сақталған екеуі 1942 ж. жиналды. Ең толық үлгі бұл болды голотип тұқымдастағы жаңа түр Мегалозавр, аталған M. wetherilli арқылы Уэллс 1954 жылы. Уэллс 1964 жылы дәл осы түрге жататын үлкен қаңқаны тапты. Оның бас сүйегінде сүйектер пайда болғанын түсініп, түрді жаңа түрге жатқызды. Дилофозавр 1970 ж Dilophosaurus wetherilli. Тұқым атауы «екі қабатты кесіртке» дегенді білдіреді және түрдің атауы Джон Ветериллді, а Навахо кеңесші. Содан кейін одан кейінгі үлгілер, оның ішінде нәресте табылды. Аяқтың іздері де жануарға, оның ішінде тыныштық іздеріне жатқызылған. Басқа түр, Dilophosaurus sinensis Қытайдан, 1993 жылы аталған, бірақ кейінірек бұл түрге жататындығы анықталды Синозавр. Ол ретінде белгіленді мемлекеттік динозавр туралы Коннектикут сол жерде табылған тректерге негізделген.
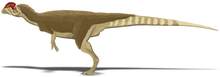
Ұзындығы 7 м (23 фут), салмағы 400 кг (880 фунт), Дилофозавр ертедегі жыртқыш динозаврлардың бірі және сол кездегі Солтүстік Америкадағы ең ірі құрлық-жануар болған. Ол жіңішке және жеңіл салынған, ал бас сүйегі пропорционалды үлкен, бірақ нәзік болатын. Тұмсығы тар, ал жоғарғы жақта мұрын тесігінен төмен саңылау немесе ирек болды. Оның бас сүйегінде бойлық, доға тәріздес жұптар болған; олардың толық пішіні белгісіз, бірақ олар ұлғайтылған болуы мүмкін кератин. The төменгі жақ сүйегі алдыңғы жағынан жіңішке және нәзік, бірақ артқы жағынан терең болды. Тістері ұзын, қисық, жіңішке және бүйірінен қысылған. Төменгі жақтағылар жоғарғы жаққа қарағанда әлдеқайда аз болды. Тістердің көпшілігінде болды серрациялар олардың алдыңғы және артқы шеттерінде. Мойын ұзын, ал омыртқалары қуыс және өте жеңіл болатын. Қолдар қуатты, ұзын және жіңішке жоғарғы сүйек сүйегімен болған. Қолдарда төрт саусақ болды; біріншісі қысқа, бірақ мықты және үлкен тырнақты көтерді, келесі екі саусақ кішірек тырнағымен ұзын және жіңішке болды; төртіншісі қалдық. Жамбас сүйегі массивті, аяқтары тік, саусақтарында үлкен тырнақтар болған.
Дилофозавр отбасы мүшесі Dilophosauridae бірге Драковенатор, арасында орналастырылған топ Coelophysidae кейінірек тероподтар пайда болды. Дилофозавр белсенді және екі аяқты болған болар еді, мүмкін ірі жануарларды аулаған болар еді; ол сондай-ақ кішігірім жануарлар мен балықтармен қоректенуі мүмкін еді. Қозғалыстың шектелуіне және алдыңғы аяқ-қолдардың қысқа болуына байланысты, аузы оның орнына жыртқышпен бірінші рет байланысқан болуы мүмкін. Жартастардың қызметі белгісіз; олар ұрыс үшін өте әлсіз болды, бірақ қолданылған болуы мүмкін визуалды дисплей, сияқты түрлерді тану және жыныстық таңдау. Ол тез өсіп, өмірінің басында жылына 30-дан 35 кг-ға дейін (66-дан 77 фунтқа дейін) өсу мүмкін. Холотип үлгісі бірнеше болды палеопатология соның ішінде емделген жарақаттар мен даму аномалиясының белгілері. Дилофозавр белгілі Kayenta формациясы сияқты динозаврлармен қатар өмір сүрді Мегапнозавр және Сарсаурус. Дилофозавр романында көрсетілген Юра паркі және оның киноны бейімдеу, онда уды түкіру және кеңейту үшін ойдан шығарылған қабілеттер берілді мойын қабыршақ, сондай-ақ нақты жануардан кішірек.
Ашылу тарихы

1942 жылдың жазында палеонтолог Чарльз Л.Кэмп бастап көшпелі кеш өткізді Калифорния университетінің Палеонтология мұражайы (UCMP) қазба қалдықтарын іздеу омыртқалылар жылы Навахо округі жылы солтүстік Аризона. Бұл туралы сөз адамдар арасында тарады Таза американдықтар сонда және Навахо Джесси Уильямс экспедицияның үш мүшесін 1940 жылы өзі тапқан сүйектердің сүйектеріне алып келді. Бұл аймақ Кайента формациясының бөлігі болды, солтүстіктен 32 км (20 миль). Кэмерон жақын Tuba City ішінде Үндістандық Навахо брондау. Күлгін түсті үш динозавр қаңқасы табылды тақтатас, үшбұрышта орналасқан, ұзындығы бір жағынан шамамен 9,1 м (30 фут). Біріншісі толығымен аяқталды, оған тек бас сүйектің алдыңғы бөлігі, жамбас бөліктері және кейбір омыртқалар жетіспеді. Екіншісі қатты эрозияға ұшырады, оған бас сүйектің алдыңғы бөлігі, төменгі жақтар, кейбір омыртқалар, аяқ-қол сүйектері және буын тәрізді қол кірді. Үшіншісі эрозияға ұшырағаны соншалық, ол тек омыртқа сынықтарынан тұрады. Бірінші жақсы онтогенез 10 күн жұмыс істегеннен кейін гипстің қаптамасына салынып, жүк машинасына тиелді, екінші онтогенез оңай жиналды, өйткені ол толығымен жердің астында қалды, бірақ үшінші онтогенезі жоғалып кетті.[2][3][4]
Палеонтологтың бақылауымен UCMP-де толықтай тазартылған алғашқы үлгі тазартылды және орнатылды Уэнн Лэнгстон, бұл процесс үш адамға екі жыл қажет болды. Қаңқа қабырғаға бекітілген бас рельеф, құйрықты жоғары қарай қисайта отырып, мойын түзетіліп, сол аяғы көріну үшін жоғары көтерілді, ал қалған қаңқа жерленген күйінде сақталды. Бас сүйегі ұсақталғандықтан, оны бірінші үлгінің бас сүйегінің артқы жағы мен екіншісінің алдыңғы бөлігі негізінде қалпына келтірді. Осыдан кейін жамбас қалпына келтірілді Аллозавр, және аяқтар да қайта қалпына келтірілді. Сол уақытта бұл теропод динозаврының толық сақталмаған қаңқаларының бірі болды. 1954 жылы палеонтолог Уэллс, онтогенезді қазған топтың бір бөлігі болған, алдын-ала бұл динозаврды қазіргі түрдегі жаңа түр ретінде сипаттап, атаған түр Мегалозавр, M. wetherilli. Толық дерлік үлгі (UCMP 37302 ретінде каталогталған) түрдің голотипі, ал екінші үлгі (UCMP 37303) жасалды паратип. The нақты атауы Уэллс «зерттеуші, ғалымдардың досы және сенімді саудагер» деп сипаттаған Навахо кеңесшісі Джон Ветериллді құрметтеді. Ветериллдің немере ағасы Милтон экспедицияға қазба қалдықтары туралы алдымен хабарлаған. Уэллс жаңа түрлерін орналастырды Мегалозавр оның ұқсас мүшелік пропорцияларына байланысты және M. bucklandiiжәне ол олардың арасында үлкен айырмашылықтар таппағандықтан. Сол уақытта, Мегалозавр ретінде қолданылғансебет таксоны », мұнда тероподтардың көптеген түрлері олардың жасына және мекеніне қарамастан орналастырылды.[2][5][3][6]

Уэллс 1964 жылы Кайба формациясының жасын анықтау үшін Туба қаласына оралды (бұл ұсынылған болатын) Кейінгі триас жасы бойынша, ал Уэллстің ойынша Ерте дейін Орта юра ) және 400 м-ге жуық басқа қаңқа тапты (1⁄4 1942 жылғы үлгілер табылған жерден оңтүстікке қарай. Толық дерлік үлгі (UCMP 77270 ретінде каталогталған) Уильям Бридтің көмегімен жиналды Солтүстік Аризона мұражайы және басқалар. Осы үлгіні дайындау кезінде оның үлкенірек адам екендігі белгілі болды M. wetherilliжәне оның бас сүйегінің жоғарғы жағында екі дөңес болған болар еді. Жіңішке сүйек тәрізді тақта болғандықтан, бір шың басынан сүйегінің сол жақ бөлігінің бір бөлігі деп саналды, оны өз орнынан жұлып алған қоқыс. Оның шыңы екендігі анықталған кезде, сонымен бірге тиісті шыңның сол жағында болатындығы түсінілді, өйткені оң жотасы орта сызықтан оң жақта және орта ұзындығы бойынша ойыс болған. Бұл жаңалық голотип үлгісін қайта тексеруге алып келді, оның екі ұсақ, жоғары қарай созылған сүйектерінің негіздері бар екендігі анықталды, олар бірге ұсақталды. Бұл бұдырлар да бейнеленген, бірақ олар бұрын дұрыс емес сүйек сүйектерінің бөлігі болып саналған. 1942 жылғы екі үлгі де табылды кәмелетке толмағандар, 1964 жылғы үлгі ересек адам болған, ал басқаларына қарағанда шамамен үштен бірі үлкен.[2][7][8] Уэллс кейінірек кресттерді «құртқа қанат» табу сияқты күтпеген нәрсе деп ойлағанын еске түсірді.[9]

Уэллс пен көмекші жаңа қаңқаға сүйене отырып, голотип үлгісінің қабырғадағы тіреуіштерін кресттерді қалпына келтіру, жамбас сүйектерін қалпына келтіру, мойын қабырғаларын ұзарту және оларды бір-біріне жақын орналастыру арқылы түзетті. Солтүстік Америка және Еуропа тероподтарының қаңқаларын зерттегеннен кейін Уэллс динозаврға тиесілі емес екенін түсінді Мегалозавржәне жаңа тұқым атауы қажет болды. Ол кезде бастарында үлкен бойлық қыртыстары бар басқа тероподтар белгілі болған жоқ, сондықтан динозавр палеонтологтардың қызығушылығына ие болды. Холотип үлгісінің формасы жасалып, оның әйнектен жасалған шыны талшықтары әртүрлі экспонаттарға таратылды; осы кастингтерді таңбалауды жеңілдету үшін Уэллс жаңа түрді егжей-тегжейлі сипаттама жарияланғанға дейін күткеннен гөрі қысқаша ескертуде атауға шешім қабылдады. 1970 жылы Уэллс жаңа түрдің атын ұсынды Дилофозавр, грек сөздерінен алынған ди (δι) «екі» мағынасын, лофос (λόφος) «шың», және мағынасын білдіреді саурос (σαυρος) «кесіртке» мағынасын білдіреді: «екі қабатты кесіртке». Уэллс егжей-тегжейлі жариялады остеологиялық сипаттамасы Дилофозавр 1984 жылы, бірақ 1964 үлгісін қоспады, өйткені ол басқа түрге жатады деп ойлады.[2][7][10][8][11] Дилофозавр ерте юра дәуірінен белгілі алғашқы теропод болды және сол дәуірдегі ең жақсы сақталған мысалдардың бірі болып қала береді.[5]
2001 жылы палеонтолог Роберт Дж. Гей кем дегенде үшеуінің қалдықтарын анықтады Дилофозавр үлгілері (бұл сан үш аралық сүйектің сынықтары мен екі түрлі өлшемді фемораның болуына негізделген) Солтүстік Аризона мұражайының коллекцияларында. Үлгілер 1978 жылы «Үлкен теропод» деп таңбаланған 190 км (120 миль) қашықтықтағы Рок-Хадраджде табылды. Материалдың көп бөлігі зақымдалған болса да, жамбастың бөлігі мен бірнеше қабырғаны қоса алғанда, алдыңғы үлгілерде сақталмаған элементтерді қосқанда маңызды. Коллекцияның кейбір элементтері осы тұқымның ең жас үлгісі және Солтүстік Америкадан шыққан ең ерте жастағы тероподтардың бірі болып табылатын нәресте үлгісіне (MNA P1.3181) тиесілі болды, олардың алдынан тек кейбіреулер шыққан Цеелофиз үлгілер. Кәмелетке толмаған үлгіні ішінара иық сүйегі, жартылай фибула және тіс сынықтары құрайды.[12] 2005 жылы палеонтолог Рональд С. Тыкоски Аризона штатындағы Голд-Спрингтен (TMM 43646-140) үлгісін тағайындады. Дилофозавр, бірақ 2012 жылы палеонтолог Мэттью Т. Каррано және оның әріптестері мұның кейбір бөлшектерімен ерекшеленетінін анықтады.[13][14]
2020 жылы палеонтологтар Адам Д. Марш пен Тимоти Б.Роу жан-жақты қайта қарастырды Дилофозавр 1964 жылдан бері сипатталмаған UCMP 77270 үлгісін қоса алғанда, сол кездегі белгілі үлгілерге негізделген. Сонымен қатар олар бұрын берілген кейбір үлгілерді алып тастап, оларды анықтау үшін тым фрагментті деп санап, типтегі карьерді басқа жерге көшірді.[6] Сұхбатында Марш қоңырау шалды Дилофозавр «ең жақсы танымал динозавр», өйткені жануар 80 жыл бұрын табылғанына қарамастан, оны нашар түсінді. Үлкен проблема - сынамалардың алдыңғы зерттеулері қай бөліктердің түпнұсқа сүйектері екендігі және олардың қайсысы гипсте қайта қалпына келтірілгені туралы түсініксіз болды, дегенмен келесі зерттеушілерде динозаврлардың анатомиясын терең түсініп, кейінгі зерттеулерге сенуге Уэллс 1984 монографиясы ғана болды. Марш динозаврға қатысты мәселелерді анықтау үшін үлгілерді зерттеуге жеті жыл жұмсады, оның ішінде екі онжылдық бұрын оның докторы Роу тапқан екі үлгіні қоса алғанда. кеңесші.[15]
Бұрын тағайындалған түрлер

1984 жылы Уэллс 1964 жылғы үлгіге (UCMP 77270) тиесілі емес деген болжам жасады Дилофозавр, бірақ бас сүйегінің, омыртқалардың және фемораның айырмашылықтарына негізделген жаңа түрге. Ол екі тұқымның да тістер болғанын, бірақ олардың нақты пішіні белгісіз екенін айтты Дилофозавр.[2] Уэллс бұл жаңа динозаврды атағанға дейін 1997 жылы қайтыс болды, бірақ екеуі бөлек тұқым болды деген ой сол кезден бастап еленбеді немесе ұмытылды.[5] 1999 жылы әуесқой палеонтолог Стефан Пикеринг жаңа атауды жеке жариялады Дилофозавр оны жинауға көмектескен Бридтің құрметіне аталған 1964 жылғы үлгіге негізделген «селекция». Бұл а номен нудум, жарамсыз түрде жарияланған атау және Гей 2005 жылы арасында айтарлықтай айырмашылықтар жоқтығына назар аударды Д.. «селекция» және басқалары D. ветерилли үлгілер.[16][17] 2012 жылы Каррано және оның әріптестері 1964 жылғы үлгі мен голотип үлгісі арасындағы айырмашылықтарды тапты, бірақ оларды түрлерге емес, жеке адамдар арасындағы вариацияға жатқызды.[13] Палеонтологтар Кристоф Хендрикс пен Октавио Матеус 2014 жылы белгілі үлгілер екі түрді бейнелеуі мүмкін деген болжам жасады. Дилофозавр тағайындалған үлгілерді толық сипаттауды күтіп, бас сүйегінің әр түрлі ерекшеліктері мен стратиграфиялық бөлінуіне негізделген.[18] Марш пен Роу 2020 жылы тек біреу ғана бар деген қорытындыға келді таксон белгілі арасында Дилофозавр үлгілер, және олардың арасындағы айырмашылықтар олардың жетілу дәрежесі мен сақталуының әр түрлі болуына байланысты болды. Олар үлгілер арасында да айтарлықтай стратиграфиялық бөлуді таппады.[6]
Терропод қаңқасы (KMV 8701) табылды Lufeng қалыптастыру, жылы Юньнань провинциясы, Қытай, 1987 ж. Ол ұқсас Дилофозавр, примаксиланы жоғарғы жақ сүйегінен бөлетін жұп шыңдармен және саңылауымен, бірақ кейбір бөлшектерімен ерекшеленеді. Палеонтолог Шаоцзин Ху оны жаңа түрі деп атады Дилофозавр 1993 жылы, D. sinensis (грек тілінен алынған) Синай, Қытайға сілтеме жасай отырып).[19] 1998 жылы палеонтолог Мэттью С. Ламанна және оның әріптестері тапты D. sinensis бірдей болу Sinosaurus triassicus, сол формациядан шыққан теропод, 1940 ж.[20] Бұл тұжырымды палеонтолог Лида Син мен оның әріптестері 2013 жылы растады, ал палеонтолог Гуо-Фу Ванг және оның әріптестері бұл түрге жататынына келіскен Синозавр 2017 жылы олар бұл жеке түр болуы мүмкін деп болжады, S. sinensis.[21][22]
Сипаттама
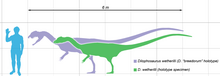
Дилофозавр алғашқы ірі жыртқыштардың бірі болды динозаврлар, орташа теропод кейбір кейінгі тероподтармен салыстырғанда аз болса да.[2][5] Бұл ерте юра дәуірінде Солтүстік Америкадағы ең ірі құрлық-жануар болған.[6] Жіңішке және жеңіл салынған, оның өлшемін а өлшемімен салыстыруға болатын қоңыр аю.[5][23][24] Белгілі ең үлкен үлгінің салмағы 400 килограмм (880 фунт), ұзындығы 7 метр (23 фут) болатын, ал оның бас сүйегі 590 миллиметр (23 1⁄4 жылы) ұзақ. Кішірек голотип үлгісі шамамен 283 килограмм (624 фунт), 6,03 метрді (19 фут) құрады 9 1⁄2 ұзындығы, жамбастың биіктігі шамамен 1,36 метр (4 фут) 5 1⁄2 және оның бас сүйегі 523 миллиметрді (1 фут) құрады 8 1⁄2 жылы) ұзақ.[23][25] Тероподтың ұқсас іздеуі Дилофозавр және Лилиенстернус кейбір зерттеушілер әсер қалдырған деп түсіндірді қауырсындар іштің және аяқтың айналасында, ұқсас төмен.[26][27] Басқа зерттеушілер оның орнына бұл әсерлерді түсіндіреді седиментологиялық динозавр қозғалған кезде жасалған артефактілер, бірақ бұл интерпретация трек-мейкердің қауырсынды болуы мүмкін екенін жоққа шығармайды.[28][29]
Бас сүйегі

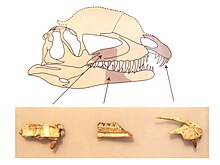
Бас сүйегі Дилофозавр жалпы қаңқаға пропорционалды үлкен болғанымен, нәзік болды. Тұмсық алдыңғы көріністе тар болып, дөңгеленген төбеге қарай тарыла түсті. The премаксилла (жоғарғы жақтың алдыңғы сүйегі) бүйірінен қарағанда ұзын және төмен, алдыңғы жағынан пиязшық тәрізді, ал оның сыртқы беті тұмсығынан нариске дейін (дөңес сүйек тесігі) аз дөңес болды. Мұрын тесіктері көптеген басқа тероподтарға қарағанда артқа қойылды. Премаксилалар бір-бірімен тығыз артикуляцияда болды, ал премаксилалар тек қана байланысты болды жоғарғы жақ сүйегі (жоғарғы жақтың келесі сүйегі) таңдайдың ортасында, бүйірінде байланыссыз, олар осы сүйектердің артқы және алға бағытталған процестері арасындағы өзара тығыз байланыстағы мықты буын құрды. Артқы және одан төмен, премаксилла субнариальды саңылау деп аталатын өзімен және жоғарғы жақ сүйектері арасындағы саңылау үшін қабырға құрды («кинк» деп те аталады). Мұндай алшақтық тағы бар целофифоидтар, сонымен қатар басқа динозаврлар. Субнариальды алшақтық а диастема, тіс қатарындағы саңылау (оны «ойық» деп те атаған). Субнариальды саңылаудың астында премаксилланың тіс тесігінің арғы жағында премаксиланың төмен қараған кильімен қоршалған субнариальды шұңқыр деп аталатын терең қазба болды.[2][23][30][7][6]
Премаксиланың сыртқы беті жабылған форамина (саңылаулар) әртүрлі мөлшерде. Премаксиланың артқа қарай созылған екі процесінің жоғарғы жағы ұзақ және төмен болды және созылған наристің жоғарғы шекарасының көп бөлігін құрады. Оның шрифті көлбеу болды, бұл аймақты базалық ойыспен профильге айналдырды. Бар премаксиланың төменгі жағы альвеолалар (тіс ұяшықтары) сопақша болды. Жоғарғы жақ сүйегі таяз және айналасында депрессияға ұшыраған antorbital fenestra (көздің алдындағы үлкен саңылау), алдыңғы жағына қарай дөңгелектелген ойық түзетін және максилардың қалған бөліктеріне қарағанда тегіс. Алдыңғы иілісте осы ойыққа преанторбитальды фенестра деп аталатын тесік ашылды. Жоғарғы жақ сүйегі бүйір жағынан альвеоладан жоғары жүгірді. Терең қоректік ойық табан бойымен субнариялық шұңқырдан артқа қарай созылды тісаралық плиталар (немесе ругозалар) жоғарғы жақ сүйектері.[2]
Дилофозавр бас сүйегінің төбесінде ұзын бойлы жұқа және доға тәрізді (немесе тәрелке тәрізді) төбешіктер болды. Шыңдар (назолакримальды шыңдар деп аталады) премаксилда төмен жоталардан басталып, негізінен жоғары қарай кеңейтілген мұрын және лакрималды сүйектер. Бұл сүйектер болды бірлескен бірге (сүйек тінін қалыптастыру кезінде бірігу), сондықтан тігістер олардың арасын анықтау мүмкін емес. Лакрималды сүйек қалың, кедір-бұдырға ұласты преорбитальды бастың жоғарғы шекарасында доға түзеді орбита (көз саңылауы), жотаның артқы жағының тіреуіші. Бұл түрге ерекше түрде орбитаның үстіндегі жиек артқа қарай жалғасып, орбитаның артында кішкене сыртқа қарай қисайған үшбұрышты дерлік процесте аяқталды. Бұл процестің үстіңгі бетінің қысқа бөлігі ғана үзілмегендіктен, қалған жотасы бас сүйегінен ~ 12 миллиметр (0,47 дюйм) қашықтыққа көтерілген болуы мүмкін. UCMP 77270 шыңының сақталған бөлігі анторбиталь фенестрасының ортаңғы нүктесінде ең биік. UCMP 77270 төбешіктердің негіздері арасындағы ойыс сөрені сақтайды және алдыңғы жағынан қарағанда олар ~ 80 ° бұрышпен жоғары және бүйір жаққа проекцияланған. Уэллс екі қабатты еске түсіретін шыңдарды тапты казуарлық Марш пен Роу олардың қамтылғанын мәлімдеді кератин немесе кератинделген тері. Олар салыстыру арқылы деп атап өтті дулыға, төбелеріндегі кератин Дилофозавр оларды сүйек көрсеткеннен гөрі көбірек үлкейте алар еді. Бір ғана үлгі кресттердің көп бөлігін сақтайтындықтан, олардың жеке адамдар арасындағы айырмашылықтары белгісіз.[6][2][7][10][5][13]
Орбита сопақша болып, түбіне қарай тар болды. The жілік сүйегі жоғары бағытталған екі процесс болды, олардың біріншісі анторбитальды фенестраның төменгі жиегінің бөлігін, ал орбитаның төменгі жиегінің бір бөлігін құрады. Проекциясы квадрат сүйегі ішіне бүйірлік уақытша фенестра (көздің артында ашу) осыны берді reniform (бүйрек тәрізді) контур. The foramen magnum (артқы жағындағы үлкен тесік бринказа ) өзі болған оксипитальды саңылаудың шамамен жартысына тең болды кордиформ (жүрек тәрізді), ал қысқа мойны және бүйірінде ойығы болған.[2] The төменгі жақ сүйегі алдыңғы жағынан жіңішке және нәзік болды, бірақ буын аймағы (бас сүйегімен байланысқан жерде) массивті болды, ал төменгі жақ сүйегі тереңде төменгі жақ фенестрасы (бүйіріндегі тесік). Төменгі жақ сүйектері кішігірім болды Дилофозавр, целофифоидтармен салыстырғанда және алдыңғы жағынан артқа қарай азайтылған, тек осы түрге арналған. The тіс сүйегі (тістердің көп бөлігі бекітілген төменгі жақ сүйегінің алдыңғы бөлігінде) сүйек емес, жоғары қисық болған. Иектің ұшында үлкен тесік болған, ал кішкене тесік қатар тіс қатарының жоғарғы жиегімен параллель параллель жүрді. Ішкі жағынан төменгі жақ симфизі (төменгі жақтың екі жартысы қосылған жерде) тегіс және тегіс болды, және оның қарама-қарсы жартысымен балқудың белгісі жоқ. A Меккелік форамен тіс қатарының сыртқы жағымен жүгірді. Бүйір беті бұрышты сүйек квадратпен артикуляция алдында ерекше пирамидалық процесті өткізді және бұл көлденең жотасы сөре құрады. The артқы төменгі жақ сүйегі (артқа проекция) ұзақ болды.[2][30][6]
Дилофозавр әр премаксиллада төрт, жоғарғы жақта 12 және әр тіс қатарында 17 тіс болған. Тістер, негізінен, ұзын, жіңішке және қайталанатын, негіздері салыстырмалы түрде кішкентай болды. Олар бүйір жағынан қысылған, көлденең қимасы бойынша сопақша, жоғарыдан линзалық (линза тәрізді), ал сыртқы және ішкі жағынан сәл вогнуты. Жоғарғы жақ сүйегінің ең үлкен тісі төртінші альвеолада немесе оның жанында болды, ал тіс тәждерінің биіктігі артқа қарай төмендеді. Жоғарғы жақ сүйегінің бірінші тісі оның альвеоласынан сәл алға қарай бағытталған, өйткені прексамилла процесінің төменгі шекарасы (максимумға қарай артқа қарай проекцияланған). Тіс сауытының тістері жоғарғы жақ сүйектеріне қарағанда әлдеқайда аз болды. Үшінші немесе төртінші тіс Дилофозавр ал кейбір целофифоидтар сол жерде ең үлкені болды және жоғарғы жақтың субнариалды саңылауына сәйкес келген сияқты. Тістердің көпшілігінің алдыңғы және артқы шеттерінде тік ойықтармен ығысқан, алдыңғы жағынан кішірек тістер пайда болды. Алдыңғы жиектерде шамамен 31-ден 41-ге дейін, ал артқы жағында 29-дан 33-ке дейін. Кем дегенде, премаксилланың екінші және үшінші тістерінде серрациялар болған, бірақ төртінші тістерде болмады. Тістері жұқа қабатпен жабылған эмаль, Қалыңдығы 0,1-ден 0,15 мм-ге дейін (0,0039-ден 0,0059 дюймге дейін), олардың негіздеріне қарай ұзарған. Альвеолалар эллипс тәрізді дөңгелек пішінді болды және олардың барлығы тістердің негіздерінен үлкен болды, сондықтан олар жақта ұсталмаған болуы мүмкін. Тіс қатарындағы альвеолалар саны тістердің өте көп екенін көрсетсе де, олардың альвеолаларының үлкендігіне байланысты олар бір-бірінен едәуір алшақ орналасқан. Құрамында иек бар ауыстырылатын тістер атқылаудың әртүрлі кезеңдерінде. Тістер арасындағы тіс аралық плиталар өте төмен болды.[2][30][12]
Посткраниялық қаңқа

Дилофозавр 10 мойны (мойны), 14 доральді (артқы жағы) және 45 каудальды (құйрық) омыртқалары болған. Оның бас сүйегі мен иығынан бас сүйегін көлденең қалыпта ұстап, шамамен 90 ° бүгілген мойны болатын. Жатыр мойны омыртқалары ерекше жеңіл болды; олардың центрлері (омыртқалардың «денелері») ойықтан шығарылды плеврокельдер (бүйіріндегі ойпаттар) және центрокельдер (ішіндегі қуыстар). Мойын омыртқаларының доғаларында да конустық ойықтардың үлкендігі, оларды бөлетін сүйектер кейде қағаз тәрізді жұқа болатын. Централар плано-вогнуты, алдыңғы жағынан тегіс-әлсіз дөңес, ал артқы жағында терең кесілген (немесе ойыс) болды. Ceratosaurus. Бұл мойынның центрада біріктірілген ұзын, мойын қабырғалары болғанымен, икемді екенін көрсетеді. Жатыр мойнының қабырғалары жіңішке болды, олар оңай бүгілген болуы мүмкін.[2][30]
The атлас сүйегі (бас сүйегіне жабысатын алғашқы мойын омыртқасы) шағын, центрлік центрі бар, ал алдыңғы жағында ойығы бар, онда ол шыныаяқ жасайтын. желке ішек (атлас омыртқасымен байланысатын өсінді) бас сүйегінің артқы жағында. The осьтік сүйек (екінші мойын омыртқасында) ауыр омыртқа болды және оның постзигафофиздер (келесі омыртқалардың презигапофиздерімен буындар жасайтын омыртқалардың процестері) үшінші мойын омыртқасынан жоғары қарай қисайған ұзын презигафофиздер кездесті. Жатыр мойны омыртқаларының центрлік және нервтік омыртқалары ұзын және төмен болды, ал омыртқалар бүйірлік көріністе басылып, алдыңғы және артқы жағында «иықтар» қалыптастырды, сондай-ақ биік, орталық «қалпақшалар» көрініс берді. Мальта кресі (крест тәрізді) жоғарыдан көрінгенде, осы динозаврдың ерекше белгілері. Жатыр мойнының артқы центродиапофизальды қабаты серияның өзгеруін, бифуркацияны және мойынға қайта қосылуын көрсетті. Жұлын омыртқаларының нервтік омыртқалары да төмен және кеңейтілген алдыңғы және артқы жағында болды, бұл мықты тіректер қалыптастырды байламдар. Бұл тұқым үшін ерекше ортаңғы діңдік омыртқалардың алдыңғы центродиапофизальды ламиналарынан және артқы центродиапофизальды ламиналардан қосымша ламиналар пайда болды. The сакральды омыртқалар ұзындығын алып жатқан ilium пышақ қосылмаған сияқты. Бірінші сакральды омыртқаның қабырға қабығы, қабықтың алдын-ала қабыну үрдісімен анықталған, бұл оның ерекшелігі. Каудальды омыртқалардың центрі ұзындыққа өте сәйкес болды, бірақ олардың диаметрі артқы жағына қарай кішірейіп, көлденең қимада эллипс тәрізді дөңгелектеніп өтті.[2][30][6]

The скапула (иық пышақтары) орташа ұзындықта және дененің қисаюын қадағалау үшін ішкі жағында ойыс болған. Скапула кең, әсіресе төртбұрышты (немесе төртбұрышты) жоғарғы бөлігі, ерекше ерекшелігі болды. The коракоидтар эллипс тәрізді болды және скапулаға қосылмаған. Коракоидтардың төменгі артқы бөліктерінде бицепс түйнегінің жанында «көлденең тірек» болды, тек осы түрге тән. Қолдар күшті болды, терең шұңқырлар мен бұлшықеттер мен байламдарды бекітуге арналған қатты процестер болды. The гумерус (қолдың жоғарғы сүйегі) үлкен және жіңішке, мықты эпиподиальды және ульна (қолдың төменгі сүйегі) тік және түзу, тікенді олекранон. Қолдарда төрт саусақ болды: біріншісі келесі екі саусаққа қарағанда қысқа, бірақ мықты, үлкен тырнақпен, ал келесі екі саусақ ұзын әрі жіңішке, кішкентай тырнақтармен. Тырнақтар қисық және өткір болды. Үшінші саусақ кішірейіп, төртіншісі кішірейген қалдық (сақталған, бірақ функциясы жоқ).[2][30][6]
The шың илиумның илиальға қарағанда ең жоғарысы болды педункул (илиумның төмен түсу процесі), ал оның сыртқы жағы ойыс болды. Аяғы ішек сүйегі сәл ғана кеңейтілген, ал төменгі жағы едәуір кеңейтілген ишкиум, оның білігі де өте жұқа болды. Артқы аяқтары үлкен болды, ұзын шлиппен сан сүйегі (жамбас сүйегі) қарағанда жіліншік (төменгі аяқтың сүйегі), мысалы, керісінше, Цеелофиз. Сан сүйегі массивті болды; оның білігі болды сигмоидты -пішінді ('S' тәрізді қисық) және оның үлкен троянтер біліктің ортасына орналастырылған. Тибия дамыған тубероздық және төменгі жағында кеңейтілді. The астрагал сүйегі (тобық сүйегі) жіліншіктен және кальцений, және фибула үшін ұяшықтың жартысын құрады. Оның қолындағыларға қарағанда әлдеқайда аз қисық, үлкен тырнақтары бар, үш дамыған саусақтары бар, ұзын аяқтары болды. Үшінші саусақ ең қатал болды, ал кіші бірінші саусақ ( hallux ) жерде ұсталмады.[2][30][31][6]
Жіктелуі

Уэллс ойлады Дилофозавр а мегалозавр 1954 жылы, бірақ 1970 жылы оның ойықтары бар екенін анықтағаннан кейін өз пікірін қайта қарады.[7][3] 1974 жылға қарай Уэллс пен палеонтолог Роберт А. Лонг тапты Дилофозавр болу цератозавроид.[32] 1984 жылы Уэллс мұны тапты Дилофозавр екеуінің де ерекшеліктерін көрсетті Коелурозаврия және Карнозаврия, дененің өлшеміне сүйене отырып, осы кезге дейін тероподтар бөлінген екі негізгі топ және ол бұл бөлудің дұрыс емес екенін болжады. Ол тапты Дилофозавр әдетте отбасында орналастырылатын тероподтарға жақын болу Halticosauridae, атап айтқанда Лилиенстернус.[2]
1988 жылы палеонтолог Григорий С.Павл галтикозаврларды отбасының субфамилиясына жатқызды Coelophysidae, және ұсынды Дилофозавр тікелей ұрпағы болуы мүмкін еді Цеелофиз. Пауыл бұл мүмкіндікті де қарастырды спинозаврлар мылжың тұмсығының, мұрынның орналасуы мен жіңішке тістерінің ұқсастығына негізделген, кейінірек тірі қалған дилофосаврлар болды. Барионекс.[23] 1994 жылы палеонтолог Томас Р. Холтц орналастырылған Дилофозавр Coelophysoidea тобында, Coelophysidae-мен бірге, бірақ бөлек. Ол Coelophysoidea-ны Ceratosauria тобына орналастырды.[33] 2000 жылы палеонтолог Джеймс Х.Мадсен мен Уэллс Цератозаврияны отбасыларға бөлді Ceratosauridae және Dilophosauridae, бірге Дилофозавр соңғы отбасының жалғыз мүшесі ретінде.[34]

Ламанна және оның әріптестері 1998 жылдан бері атап өтті Дилофозавр оның бас сүйегінде дөңестері бар екендігі анықталды, басқа тәрізді крест тәрізді тероподтар табылды (соның ішінде Синозавр), және бұл ерекшелік тек тұқымға ғана тән емес және олардың тобындағы өзара байланысты анықтау үшін шектеулі пайдалану.[20] Палеонтолог Адам М.Ятс тұқымдасты сипаттады Драковенатор 2005 жылы Оңтүстік Африкадан келіп, оны тығыз байланысты деп тапты Дилофозавр және Zupaysaurus. Оның кладистикалық талдау олар Coelophysoidea-ға жатпайды, керісінше Неотеропода, көбірек алынған (немесе «озық») топ. Егер ол ұсынды Дилофозавр Coelophysoidea-ға қарағанда көбірек алынған, оның осы топпен бөліскен ерекшеліктері мұраға қалған болуы мүмкін базальды (немесе «қарабайыр») тероподтар, бұл тероподтардың алғашқы эволюциясында «целофифоидтық сатыдан» өткен болуы мүмкін екенін көрсетеді.[35]
2007 жылы палеонтолог Натан Д.Смит және оның әріптестері крест тәрізді тероподты тапты Криолофозавр болу қарындас түрлер туралы Дилофозавржәне оларды топтастырды Драковенатор және Синозавр. Бұл жабын Coelophysoidea-дан көбірек алынған, бірақ Цератозавриядан гөрі базальды, осылайша базальды тероподтарды баспалдақ тәрізді етіп орналастырды.[36] 2012 жылы Каррано және оның әріптестері Смит пен оның әріптестері ұсынған крест тәрізді тероподтар тобы осындай кресттердің болуына қатысты ерекшеліктерге негізделген, бірақ онтогенездің қалған бөліктерінің ерекшеліктері онша сәйкес келмейтіндігін анықтады. Олар мұны тапты Дилофозавр целофифоидты болды Криолофозавр және Синозавр топтың базальды мүшелері ретінде көбірек алынған Тетанурае.[13]
Палеонтолог Кристоф Хендриккс және оның әріптестері Dilophosauridae-ді қосуды анықтады Дилофозавр және Драковенатор 2015 жылы және осы топты орналастыруға қатысты жалпы белгісіздік болғанымен, ол Coelophysoidea-ға қарағанда әлдеқайда көбірек пайда болатын көрінеді, ал қарындас топ Аверостра. Dilophosauridae Coelophysoidea-мен субнариальды саңылау және жоғарғы жақ сүйектерінің алдыңғы тістері сияқты бөліседі, ал Аверостраға ортақ ерекшеліктерге максилардың алдыңғы бөлігіндегі фенестра және жоғарғы жақ тістерінің саны азаяды. Олар краниальды кресттерді ұсынды Криолофозавр және Синозавр екеуі де болды конвергентті түрде дамыды, немесе ортақ атадан мирас болған ерекшелік болды. Келесісі кладограмма Хендриккс және оның әріптестері, бұрын жүргізілген зерттеулерге негізделген:[37]
| Неотеропода |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
2019 жылы палеонтологтар Марион Захнер мен Винанд Бринкманн Dilophosauridae мүшелерін Авеространың дәйекті базальды таксондары деп тапты монофилетикалық клад (табиғи топ), бірақ олардың кейбір талдаулары құрамында топтың бар екендігін анықтады Дилофозавр, Драковенатор, Криолофозавр, және мүмкін Notatesseraeraptor ең негізгі мүше ретінде. Сондықтан олар Dilophosauridae-ге төменгі жақтың ерекшеліктеріне сүйене отырып диагноз қойды.[38] Марш пен Роу оларды 2020 жылы қайта шығарумен бірге жүретін филогенетикалық анализде барлық үлгілерді тапты Дилофозавр монофилетикалық топ құру, Аверостраға қарындас және одан да көп туынды Криолофозавр. Олардың талдаулары Dilophosauridae-ге қолдау таппады және олар ми сүйектері a плезиоморфты цератозаврия мен тетанураның (тектік) қасиеті.[6]
Ихнология

Әр түрлі ichnotaxa (таксондар негізделген қазба қалдықтарын іздеу ) қатысты болды Дилофозавр немесе ұқсас тероподтар. 1971 жылы Уэллс солтүстіктегі Аризонаның Кайента түзілуінен 14 м (45 фут) және 112 м (367 фут) төменде екі деңгейде динозаврлардың іздері туралы хабарлады. Дилофозавр үлгілері табылды. Төменгі іздер болды тридактил (үш саусақты), және оны жасауы мүмкін еді Дилофозавр; Уэллс жаңа ішногенді және түрлерді жасады Dilophosauripus williamsi солардың негізінде, алғашқылардың ашушысы Уильямстың құрметіне Дилофозавр қаңқалар. Үлгі үлгісі - бұл UCMP 79690-4 каталогталған үлкен із іздері, гиподигмаға тағы үш баспа іздері енгізілген.[39] 1984 жылы Уэллс аяқ іздері тиесілі екенін дәлелдейтін немесе жоққа шығаратын жол табылмағанын мойындады Дилофозавр.[2] 1996 жылы палеонтологтар Майкл Моралес пен Скотт Булки а жол ішногеннің Eubrontes өте үлкен теропод жасаған Кайента формациясынан. Олар мұны өте үлкен көлемде жасауға болатындығын атап өтті Дилофозавр жеке, бірақ екіталай екенін анықтады, өйткені олар трекшінің 2,83–2,99 м болатындығын болжады (9 фут) 3 1⁄2 9 футтан кейін 9 3⁄4 1.50-1.75 м (4 фут 11 дюйм - 5 фут 9 дюйм) салыстырғанда, жамбаста биік Дилофозавр.[40]

Палеонтолог Жерар Джерлиńски тридактил іздерін зерттеді Қасиетті Крест таулары жылы Польша және олар 1991 жылы тәрізді тероподқа жатады деген қорытынды жасады Дилофозавр. Ол жаңа ішнеспецификацияларды атады Граллатор (Eubrontes) солтиковенсис олардың негізінде голотип ретінде MGIW 1560.11.12 ізі бар.[41] 1994 жылы Джерлиńски де із қалдырды Хоганас формациясы 1974 жылы Швецияда ашылды G. (E.) солтиковенсис.[42] 1996 жылы Джерлиńски AC 1/7 трассасын Тернерлер Falls Formation Массачусетс штатында, ол тероподқа ұқсас қауырсындар әсерін көрсетеді деп сенді Дилофозавр және Лилиенстернусжәне оны ичнотаксонға тағайындады Grallator minisculus.[26] Палеонтолог Мартин Кундрат тректің 2004 жылы қауырсындардан алған әсерлерін көрсетті деген пікірге келісті, бірақ палеонтолог бұл интерпретацияны даулады Мартин Локли және 2003 жылғы әріптестер және палеонтолог Энтони Дж. Мартин және оларды 2004 жылы седиментологиялық артефакт ретінде қарастырған әріптестер. Мартин және оның әріптестері тректі ішнотаксонға қайта тағайындады Fulicopus lyellii.[27][28][29]
Палеонтолог Роберт Э.Вимс 2003 жылы бұл туралы айтқан болатын Eubrontes тректерді теропод емес, а сауроподоморф ұқсас Платеозавр, қоспағанда Дилофозавр мүмкін трекмейкер ретінде. Оның орнына Weems ұсынды Kayentapus hopii, Уэллс 1971 жылы тағы бір ішнотаксонды ең жақсы матч деп атады Дилофозавр.[43] Атрибуциясы Дилофозавр was primarily based on the wide angle between digit impressions three and four shown by these tracks, and the observation that the foot of the holotype specimen shows a similarly splayed-out fourth digit. Also in 2003, paleontologist Emma Rainforth argued that the splay in the holotype foot was merely the result of distortion, and that Eubrontes would indeed be a good match for Дилофозавр.[44][45]The paleontologist Спенсер Г. Лукас and colleagues stated in 2006 that virtually universal agreement existed that Eubrontes tracks were made by a theropod like Дилофозавр, and that they and other researchers dismissed Weems' claims.[46]
In 2006, Weems defended his 2003 assessment of Eubrontes, and proposed an animal like Дилофозавр as the possible trackmaker of numerous Kayentapus trackways of the Culpeper Quarry in Virginia. Weems suggested rounded impressions associated with some of these trackways to represent hand impressions lacking digit traces, which he interpreted as a trace of quadrupedal movement.[45] Milner and colleagues used the жаңа комбинация Kayentapus soltykovensis in 2009, and suggested that Dilophosauripus may not be distinct from Eubrontes және Kayentapus. They suggested that the long claw marks that were used to distinguish Dilophosauripus may be an artifact of dragging. They found that Gigandipus және Анхизаврипус tracks may likewise also just represent variations of Eubrontes. They pointed out that differences between ichnotaxa may reflect how the trackmaker interacted with the substrate rather than taxonomy. Олар сондай-ақ тапты Дилофозавр to be a suitable match for a Eubrontes trackway and resting trace (SGDS 18.T1) from the St. George dinosaur discovery site ішінде Moenave Formation of Utah, though the dinosaur itself is not known from the formation, which is slightly older than the Kayenta Formation.[47] Weems stated in 2019 that Eubrontes tracks do not reflect the gracile feet of Дилофозавр, and argued they were instead made by the bipedal sauropodopormph Анхизавр.[48]
Палеобиология

Азықтандыру және диета
Welles found that Дилофозавр did not have a powerful bite, due to weakness caused by the subnarial gap. He thought that it used its front premaxillary teeth for plucking and tearing rather than biting, and the maxillary teeth further back for piercing and slicing. He thought that it was probably a scavenger rather than a predator, and that if it did kill large animals, it would have done so with its hands and feet rather than its jaws. Welles did not find evidence of cranial kinesis бас сүйегінде Дилофозавр, a feature that allows individual bones of the skull to move in relation to each other.[2] In 1986, the paleontologist Роберт Т. Баккер instead found Дилофозавр, with its massive neck and skull and large upper teeth, to have been adapted for killing large prey, and strong enough to attack any Early Jurassic herbivores.[49] In 1988, Paul dismissed the idea that Дилофозавр was a scavenger, and claimed that strictly scavenging terrestrial animals are a myth. He stated that the snout of Дилофозавр was better braced than had been thought previously, and that the very large, slender maxillary teeth were more lethal than the claws. Paul suggested that it hunted large animals such as прозауроподтар, and that it was more capable of snapping small animals than other theropods of a similar size.[23]
2005 ж сәуле теориясы study by the palaeontologist François Therrien and colleagues found that the тістеу күші in the mandible of Дилофозавр decreased rapidly hindwards in the tooth-throw. This indicates that the front of the mandible, with its upturned chin, "розетка " of teeth, and strengthened symphysal region (similar to spinosaurids), was used to capture and manipulate prey, probably of relatively smaller size. The properties of its mandibular symphysis was similar to those of фелидтер and crocodilians that use the front of their jaws to deliver a powerful bite when subduing prey. The loads exerted on the mandibles were consistent with struggle of small prey, which may have been hunted by delivering slashing bites to wound it, and then captured with the front of the jaws after being too weakened to resist. The prey may then have been moved further back into the jaws, where the largest teeth were located, and killed by slicing bites (similar to some crocodilians) with the sideways-compressed teeth. Авторлар егер болса Дилофозавр indeed fed on small prey, possible hunting packs would have been of limited size.[50]
Milner and paleontologist Джеймс I. Киркланд suggested in 2007 that Дилофозавр had features that indicate it may have eaten fish. They pointed out that the ends of the jaws were expanded to the sides, forming a "rosette" of interlocking teeth, similar to those of spinosaurids, known to have eaten fish, and гариалдар, which is the modern қолтырауын that eats the most fish. The nasal openings were also retracted back on the jaws, similar to spinosaurids, which have even more retracted nasal openings, and this may have limited water splashing into the nostrils during fishing. Both groups also had long arms with well-developed claws, which could help when catching fish. Lake Dixie, a large lake that extended from Utah to Arizona and Nevada, would have provided abundant fish in the "post-cataclysmic", biologically more impoverished world that followed the Триас-юра жойылу оқиғасы.[51]

In 2018, Marsh and Rowe reported that the holotype specimen of the sauropodomorph Сарсаурус bore possible tooth marks scattered across the skeleton that may have been left by Дилофозавр (Syntarus was too small to have produced them) scavenging the specimen after it died (the positions of the bones may also have been disturbed by scavenging). An example of such marks can be seen on the left scapula, which has an oval depression on the surface of its upper side, and a large hole on the lower front end of the right tibia. The quarry where the holotype and paratype specimens of Сарсаурус were excavated also contained a partial immature Дилофозавр үлгі.[52] Marsh and Rowe suggested in 2020 that many of the features that distinguished Дилофозавр from earlier theropods were associated with increased body size and macropredation (preying on large animals). While Marsh and Rowe agreed that Дилофозавр could have fed on fish and small prey in the fluvial system in its environment, they pointed out that the articulation between the premaxilla and maxilla of the upper jaw was immobile and much more robust than previously thought, and that large-bodied prey could have been grasped and manipulated with the forelimbs during predation and scavenging. They considered the large bite marks on Сарсаурус specimens alongside shed teeth and the presence of a Дилофозавр specimen within the same quarry as support for this idea.[6]
Қозғалыс

Welles envisioned Дилофозавр as an active, clearly bipedal animal, similar to an enlarged түйеқұс. He found the forelimbs to have been powerful weapons, strong and flexible, and not used for locomotion. He noted that the hands were capable of grasping and slashing, of meeting each other, and reaching two-thirds up the neck. He proposed that in a sitting posture, the animal would rest on the large "foot" of its ischium, as well as its tail and feet.[2] In 1990, paleontologists Stephen and Sylvia Czerkas suggested that the weak pelvis of Дилофозавр could have been an adaptation for an aquatic lifestyle, where the water would help support its weight, and that it could have been an efficient swimmer. They found it doubtful that it would have been restricted to a watery environment, though, due to the strength and proportions of its hind limbs, which would have made it fleet-footed and agile during bipedal locomotion.[53] Paul depicted Дилофозавр bouncing on its tail while lashing out at an enemy, similar to a кенгуру.[54]
In 2005, paleontologists Phil Senter and James H. Robins examined the range of motion in the fore limbs of Дилофозавр және басқа тероподтар. They found that Дилофозавр would have been able to draw its humerus backwards until it was almost parallel with the scapula, but could not move it forwards to a more than vertical orientation. The elbow could approach full extension and flexion at a right angle, but not achieve it completely. The fingers do not appear to have been voluntarily hyperextensible (able to extend backwards, beyond their normal range), but they may have been passively hyperextensible, to resist dislocation during violent movements by captured prey.[55] A 2015 article by Senter and Robins gave recommendations for how to reconstruct the fore limb posture in bipedal dinosaurs, based on examination of various taxa, including Дилофозавр. The scapulae were held very horizontally, the resting orientation of the elbow would have been close to a right angle, and the orientation of the hand would not have deviated much from that of the lower arm.[56]

In 2018, Senter and Corwin Sullivan examined the range of motion in the fore limb joints of Дилофозавр by manipulating the bones, to test hypothesized functions of the fore limbs. They also took into account that experiments with аллигатор carcasses show that the range of motion is greater in elbows covered in soft tissue (such as шеміршек, ligaments, and muscles) than what would be indicated by manipulation of bare bones. They found that the humerus of Дилофозавр could be retracted into a position that was almost parallel with the scapula, protracted to an almost vertical level, and elevated 65°. The elbow could not be flexed past a right angle to the humerus. Қоңырау және супинация of the wrists (crossing the радиусы and ulna bones of the lower arm to turn the hand) was prevented by the radius and ulna joints not being able to roll, and the palms, therefore, faced medially, towards each other. The inability to pronate the wrists was an ancestral feature shared by theropods and other dinosaur groups. The wrist had limited mobility, and the fingers diverged during flexion, and were very hyperextensible.[57]
Senter and Sullivan concluded that Дилофозавр was able to grip and hold objects between two hands, to grip and hold small objects in one hand, to seize objects close beneath the chest, to bring an object to the mouth, to perform a display by swinging the arms in an arc along the sides of the ribcage, to scratch the chest, belly, or the half of the other fore limb farthest from the body, to seize prey beneath the chest or the base of the neck, and to clutch objects to the chest. Дилофозавр was unable to perform scratch-digging, hook-pulling, to hold objects between two fingertips of one hand, to maintain balance by extending the arms outwards to the sides, or to probe small crevices like the modern aye aye жасайды. The hyperexensility of the fingers may have prevented the prey's violent struggle from dislocating them, since it would have allowed greater motion of the fingers (with no importance to locomotion). The limited mobility of the shoulder and shortness of the fore limbs indicates that the mouth made first contact with the prey rather than the hands. Capture of prey with the fore limbs would only be possible for seizing animals small enough to fit beneath the chest of Дилофозавр, or larger prey that had been forced down with its mouth. The great length of the head and neck would have enabled the snout to extend much further than the hands.[57]


The Dilophosauripus footprints reported by Welles in 1971 were all on the same level, and were described as a "chicken yard hodge-podge" of footprints, with few forming a trackway. The footprints had been imprinted in mud, which allowed the feet to sink down 5–10 cm (2–4 in). The prints were sloppy, and the varying breadth of the toe prints indicates that mud had clung to the feet. The impressions varied according to differences in the substrate and the manner in which they were made; sometimes, the foot was planted directly, but often a backwards or forwards slip occurred as the foot came down. The positions and angles of the toes also varied considerably, which indicate they must have been quite flexible. The Dilophosauripus footprints had an offset second toe with a thick base, and very long, straight claws that were in line with the axes of the toe pads. One of the footprints was missing the claw of the second toe, perhaps due to injury.[39] In 1984, Welles interpreted the fact that three individuals were found closely together, and the presence of criss-crossed trackways nearby, as indications that Дилофозавр traveled in groups.[2] Gay agreed that they may have traveled in small groups, but noted that no direct evidence supported this, and that тасқын су could have picked up scattered bones from different individuals and deposited them together.[12]
Milner and colleagues examined the possible Дилофозавр trackway SGDS 18.T1 in 2009, which consists of typical footprints with tail drags and a more unusual resting trace, deposited in лакустрин жағажай құмтас. The trackway began with the animal first oriented approximately in parallel with the shoreline, and then stopping by a берма with both feet in parallel, whereafter it lowered its body, and brought its метатарсальдар және каллостық around its ischium to the ground; this created impressions of symmetrical "heels" and circular impressions of the ischium. The part of the tail closest to the body was kept off the ground, whereas the end further away from the body made contact with the ground. The fact that the animal rested on a slope is what enabled it to bring both hands to the ground close to the feet. After resting, the dinosaur shuffled forwards, and left new impressions with its feet, metatarsals, and ischium, but not the hands. The right foot now stepped on the print of the right hand, and the second claw of the left foot made a drag mark from the first resting position to the next. After some time, the animal stood up and moved forwards, with the left foot first, and once fully erect, it walked across the rest of the exposed surface, while leaving thin drag marks with the end of the tail.[47]
Crouching is a rarely captured behavior of theropods, and SGDS 18.T1 is the only such track with unambiguous impressions of theropod hands, which provides valuable information about how they used their forelimbs. The crouching posture was found to be very similar to that of modern birds, and shows that early theropods held the palms of their hands facing medially, towards each other. As such a posture therefore evolved early in the lineage, it may have characterized all theropods.Theropods are often depicted with their palms facing downwards, but studies of their functional anatomy have shown that they, like birds, were unable to pronate or supinate their arms. The track showed that the legs were held symmetrically with the body weight distributed between the feet and the metatarsals, which is also a feature seen in birds such as рититтер. Milner and colleagues also dismissed the idea that the Kayentapus minor track reported by Weems showed a palm imprint made by a quadrupedally walking theropod. Weems had proposed the trackmaker would have been able to move quadrupedally when walking slowly, while the digits would have been habitually hyperextended so only the palms touched the ground. Milner and colleagues found the inferred pose unnecessary, and suggested the track was instead made in a similar way as SGDS 18.T1, but without leaving traces of the digits.[47]
Crest функциясы

Welles conceded that suggestions as to the function of the crests of Дилофозавр were conjectural, but thought that, though the crests had no grooves to indicate vascularization, they could have been used for терморегуляция. He also suggested they could have been used for түрлерді тану немесе ою-өрнек.[2]The Czerkas pointed out that the crests could not have been used during battle, as their delicate structure would have been easily damaged. They suggested that they were a visual display for attracting a mate, and even thermoregulation.[53] In 1990, paleontologist Walter P. Coombs stated that the crests may have been enhanced by colors for use in display.[58]
In 2011 the paleontologists Кевин Падиан және Джон Р.Хорнер proposed that "bizarre structures" in dinosaurs in general (including crests, frills, horns, and domes) were primarily used for species recognition, and dismissed other explanations as unsupported by evidence. They noted that too few specimens of cranially ornamented theropods, including Дилофозавр, were known to test their evolutionary function statistically, and whether they represented жыныстық диморфизм немесе жыныстық жетілу.[59] In a response to Padian and Horner the same year, the paleontologists Rob J. Knell and Сэмпсон Д. argued that species recognition was not unlikely as a secondary function for "bizarre structures" in dinosaurs, but that жыныстық таңдау (used in display or combat to compete for mates) was a more likely explanation, due to the high cost of developing them, and because such structures appear to be highly variable within species.[60]
In 2013, paleontologists David E. Hone and Даррен Найш criticized the "species recognition hypothesis", and argued that no extant animals use such structures primarily for species recognition, and that Padian and Horner had ignored the possibility of mutual sexual selection (where both sexes are ornamented).[61] Marsh and Rowe agreed in 2020 that the crests of Дилофозавр likely had a role in species identification or intersexual/intrasexual selection, as in some modern birds.[6]
Даму

Welles originally interpreted the smaller Дилофозавр specimens as juveniles, and the larger specimen as an adult, later interpreting them as different species.[2][7] Paul suggested that the differences between the specimens was perhaps due to sexual dimorphism, as was seemingly also apparent in Цеелофиз, which had "robust" and "gracile" forms of the same size, that might otherwise have been regarded as separate species. Following this scheme, the smaller Дилофозавр specimen would represent a "gracile" example.[23]
In 2005 Tykoski found that most Дилофозавр specimens known were juvenile individuals, with only the largest an adult, based on the level of coossification of the bones.[14] In 2005 Gay found no evidence of the sexual dimorphism suggested by Paul (but supposedly present in Цеелофиз), and attributed the variation seen between Дилофозавр specimens to individual variation and онтогенез (changes during growth). There was no dimorphism in the skeletons, but he did not rule out that there could have been in the crests; more data was needed to determine this.[16] Based on the tiny nasal crests on a juvenile specimen, Yates had tentatively assigned to the related genus Драковенатор, he suggested that these would have grown larger as the animal became adult.[35]
The paleontologist J.S. Tkach reported a гистологиялық study (microscopical study of internal features) of Дилофозавр in 1996, conducted by taking жіңішке кесінділер of long bones and ribs of specimen UCMP 37303 (the lesser preserved of the two original skeletons). The bone tissues were well vascularized and had a fibro-lamellar structure similar to that found in other theropods and the sauropodomorph Массоспондилус. The plexiform (woven) structure of the bones suggested rapid growth, and Дилофозавр may have attained a growth rate of 30 to 35 kilograms (66 to 77 lb) per year early in life.[62]
Welles found that the replacement teeth of Дилофозавр and other theropods originated deep inside the bone, decreasing in size the farther they were from the alveolar border. There were usually two or three replacement teeth in the alveoli, with the youngest being a small, hollow тәж. The replacement teeth erupted on the outer side of the old teeth. When a tooth neared the сағыз сызығы, the inner wall between the interdental plates was resorbed and formed a nutrient notch. As the new tooth erupted, it moved outwards to center itself in the alveolus, and the nutrient notch closed over.[2]
Палеопатология

Welles noted various paleopathologies (ancient signs of disease, such as injuries and malformations) in Дилофозавр. The holotype had a сулькус (groove or furrow) on the neural arch of a cervical vertebra that may have been due to an injury or crushing, and two pits on the right humerus that may have been абсцесс (жинақтар ірің ) or artifacts. Welles also noted that it had a smaller and more delicate left humerus than the right, but with the reverse condition in its forearms. 2001 жылы палеонтолог Ральф Молнар suggested that this was caused by a developmental anomaly called құбылмалы асимметрия. This anomaly can be caused by stress in animal populations, for example due to disturbances in their environment, and may indicate more intense selective pressure. Asymmetry can also result from traumatic events in early development of an animal, which would be more randomly distributed in time.[2][63] A 2001 study conducted by paleontologist Bruce Rothschild and colleagues examined 60 Дилофозавр foot bones for signs of стресс сынықтары (which are caused by strenuous, repetitive actions), but none were found. Such injuries can be the result of very active, predatory lifestyles.[64]
In 2016 Senter and Sara L. Juengst examined the paleopathologies of the holotype specimen and found that it bore the greatest and most varied number of such maladies on the pectoral girdle and forelimb of any theropod dinosaur so far described, some of which are not known from any other dinosaur. Only six other theropods are known with more than one paleopathology on the pectoral girdle and forelimbs. The holotype specimen had eight afflicted bones, whereas no other theropod specimen is known with more than four. On its left side it had a fractured scapula and radius, and fibriscesses (like abscesses) in the ulna and the outer фаланг сүйегі бас бармақтың On the right side it had torsion of its humeral shaft, three bony tumors on its radius, a truncated articular surface of its third метакарпальды сүйек, and deformities on the first phalanx bone of the third finger. This finger was permanently deformed and unable to flex. The deformities of the humerus and the third finger may have been due to osteodysplasia, which had not been reported from non-avian dinosaurs before, but is known in birds. Affecting juvenile birds that have experienced malnutrition, this disease can cause pain in one limb, which makes the birds prefer to use the other limb instead, which thereby develops torsion.[65]

The number of traumatic events that led to these features is not certain, and it is possible that they were all caused by a single encounter, for example by crashing into a tree or rock during a fight with another animal, which may have caused puncture wounds with its claws. Since all the injuries had healed, it is certain that the Дилофозавр survived for a long time after these events, for months, perhaps years. The use of the forelimbs for prey capture must have been compromised during the healing process. The dinosaur may therefore have endured a long period of fasting or subsisted on prey that was small enough for it to dispatch with the mouth and feet, or with one forelimb. According to Senter and Juengst, the high degree of pain the dinosaur might have experienced in multiple locations for long durations also shows that it was a hardy animal. They noted that paleopathologies in dinosaurs are underreported, and that even though Welles had thoroughly described the holotype, he had mentioned only one of the pathologies found by them. They suggested that such features may sometimes be omitted because descriptions of species are concerned with their characteristics rather than abnormalities, or because such features are difficult to recognize.[65] Senter and Sullivan found that the pathologies significantly altered the range of motion in the right shoulder and right third finger of the holotype, and that estimates for range of motion may therefore not match those made for a healthy forelimb.[57]
Палеоэкология

Дилофозавр is known from the Kayenta Formation, which dates to the Синемурия және Плиенсбачиан stages of the Early Jurassic, approximately 196–183 million years ago.[66] The Kayenta Formation is part of the Glen Canyon тобы that includes formations in northern Arizona, parts of southeastern Utah, western Colorado, and northwestern New Mexico. It is composed mostly of two фация, one dominated by алевролит deposition and the other by sandstone. The siltstone facies is found in much of Arizona, while the sandstone facies is present in areas of northern Arizona, southern Utah, western Colorado, and northwestern New Mexico. The formation was primarily deposited by rivers, with the siltstone facies as the slower, more sluggish part of the river system. Kayenta Formation deposition was ended by the encroaching dune field that would become the Навахо құмтасы.[67] Нақты радиометриялық танысу of this formation has not yet been made, and the available stratigraphic correlation has been based on a combination of radiometric dates from vertebrate fossils, магнетостратиграфия, and pollen evidence.[66] Дилофозавр appears to have survived for a considerable span of time, based on the position of the specimens within the Kayenta Formation.[6]
The Kayenta Formation has yielded a small but growing assemblage of organisms. Most fossils are from the siltstone facies.[68] Most organisms known so far are vertebrates. Non-vertebrates include microbial or "algal" limestone,[69] тасталған ағаш,[70] plant impressions,[71] freshwater bivalves and snails,[67] остракодтар,[72] және омыртқасыздар қазба қалдықтарын іздеу.[69] Vertebrates are known from both body fossils and trace fossils. Vertebrates known from body fossils include[68] hybodont sharks, indeterminate сүйекті балық, өкпе балықтары,[70] salamanders,[73] the frog Prosalirus, цесилиан Eocaecilia, тасбақа Kayentachelys, а sphenodontian reptile, lizards,[74] and several early крокодиломорфтар оның ішінде Калсоясучус, Eopneumatosuchus, Kayentasuchus, және Протосух, және птерозавр Rhamphinion. Басқа Дилофозавр, several dinosaurs are known, including the theropods Мегапнозавр,[14] және Kayentavenator,[75] the sauropodomorph Сарсаурус,[76] а гетеродонтозавр, және тиреофоран Скутеллозавр. Синапсидтер қамтиды tritylodontids Диннебитодон, Кайентатериум, және Олигокиф, morganucodontids,[74] the possible early true mammal Диннетериум және а харамийид сүтқоректілер. The majority of these finds come from the vicinity of Gold Spring, Arizona.[68] Vertebrate trace fossils include копролиттер and the tracks of терапидтер, lizard-like animals, and several types of dinosaur.[69][77]
Тафономия
Welles outlined the taphonomy of the original specimens, changes that happened during their decay and fossilization. The holotype skeleton was found lying on its right side, and its head and neck were recurved – curved backwards – in the "death pose " in which dinosaur skeletons are often found. This pose was thought to be опистотонус (due to death-spasms) at the time, but may instead have been the result of how a carcass was embedded in шөгінділер. The back was straight, and the hindmost dorsal vertebrae were turned on their left sides. The caudal vertebrae extended irregularly from the pelvis, and the legs were articulated, with little displacement. Welles concluded that the specimens were buried at the place of their deaths, without having been transported much, but that the holotype specimen appears to have been disturbed by scavengers, indicated by the rotated dorsal vertebrae and crushed skull.[2][78] Gay noted that the specimens he described in 2001 showed evidence of having been transported by a stream. As none of the specimens were complete, they may have been transported over some distance, or have lain on the surface and weathered for some time before transport. They may have been transported by a су тасқыны, as indicated by the variety of animals found as fragments and bone breakage.[12]
Мәдени маңызы

Дилофозавр was featured in the 1990 novel Юра паркі, жазушының Майкл Крихтон, and its 1993 киноны бейімдеу режиссермен Стивен Спилберг. The Дилофозавр туралы Юра паркі was acknowledged as the "only serious departure from scientific veracity" in the movie's жасау book, and as the "most fictionalized" of the movie's dinosaurs in a book about Stan Winston Studios жасаған аниматроника әсерлер. For the novel, Crichton invented the dinosaur's ability to spit venom (explaining how it was able to kill prey, in spite of its seemingly weak jaws). The art department added another feature, a қорап folded against its neck that expanded and vibrated as the animal prepared to attack, similar to that of the frill-necked lizard. Шатаспас үшін Velociraptor as featured in the movie, Дилофозавр was presented as only 1.2 meters (4 ft) tall, instead of its assumed true height of about 3.0 meters (10 ft). Nicknamed "the spitter", the Дилофозавр of the movie was realized through puppeteering, and required a full body with three interchangeable heads to produce the actions required by the script. Separate legs were also constructed for a shot where the dinosaur hops by. Unlike most of the other dinosaurs in the movie, no компьютерлік кескіндер was employed when showing the Дилофозавр.[79][80][81]
The geologist J. Bret Bennington noted in 1996 that though Дилофозавр probably did not have a frill and could not spit venom like in the movie, its bite could have been venomous, as has been claimed for the Комодо айдаһары. He found that adding venom to the dinosaur was no less allowable than giving a color to its skin, which is also unknown. If the dinosaur had a frill, there would have been evidence for this in the bones, in the shape of a rigid structure to hold up the frill, or markings at the places where the muscles used to move it were attached. He also added that if it did have a frill, it would not have used it to intimidate its meal, but rather a competitor (he speculated it may have responded to a character in the movie pulling a hood over his head).[82] In a 1997 review of a book about the science of Юра паркі, the paleontologist Питер Додсон likewise pointed out the wrong scale of the film's Дилофозавр, as well as the improbability of its venom and frill.[83] Bakker pointed out in 2014 that the movie's Дилофозавр lacked the prominent notch in the upper jaw, and concluded that the movie-makers had done a good job at creating a frightening химера of different animals, but warned it could not be used to teach about the real animal.[84] Welles himself was "thrilled" to see Дилофозавр жылы Юра паркі: He noted the inaccuracies, but found them minor points, enjoyed the movie, and was happy to find the dinosaur "an internationally known actor".[85]

Сәйкес Navajo myth, the carcasses of slain monsters were "beaten into the earth", but were impossible to obliterate, and fossils have traditionally been interpreted as their remains. While Navajo people have helped paleontologists locate fossils since the 19th century, traditional beliefs suggest that the ghosts of the monsters remain in their partially buried corpses, and have to be kept there through potent rituals. Likewise, some worry that the bones of their relatives would be dug up along with dinosaur remains, and that removing fossils shows disrespect to the past lives of these beings.[86] In 2005, the historian Адриен мэрі stated Welles had noted that during the original excavation of Дилофозавр, the Navajo Williams disappeared from the excavation after some days, and speculated this was because Williams found the detailed work with fine brushes "beneath his dignity". Mayor instead pointed out that Navajo men do occupy themselves with detailed work, such as jewellery and painting, and that the explanation for Williams' departure may instead have been traditional anxiety as the skeletons emerged and were disturbed. Mayor also pointed to an incident in the 1940s when a Navajo man helped excavate a Pentaceratops skeleton as long as he did not have to touch the bones, but left the site when only a few inches of dirt were left covering them.[86] In a 1994 book, Welles said Williams had come back some days later with two Navajo women saying "that's no man's work, that's squaw's work".[9]
The cliffs in Arizona that contained the bones of Дилофозавр сонымен қатар бар петроглифтер арқылы ата-баба Пуэблоан carved onto them, and the criss-crossing tracks of the area are called Naasho’illbahitsho Biikee by the Navajo, meaning "big lizard tracks". According to Mayor, Navajos used to hold ceremonies and make offerings to these monster tracks. Tridactyl tracks were also featured as decorations on the costumes and рок-арт туралы Хопи және Зуни, probably influenced by such dinosaur tracks.[86] 2017 жылы Дилофозавр ретінде белгіленді мемлекеттік динозавр АҚШ штатының Коннектикут, to become official with the new state budget in 2019. Дилофозавр was chosen because tracks thought to have been made by similar dinosaurs were discovered in Рокки Хилл in 1966, during excavation for the Interstate Highway 91. The six tracks were assigned to the ішноспециттер Eubrontes giganteus, which was made the state fossil of Connecticut in 1991. The area they were found in had been a Triassic lake, and when the significance of the area was confirmed, the highway was rerouted, and the area made a мемлекеттік саябақ аталған Динозавр мемлекеттік паркі. In 1981 a sculpture of Дилофозавр, the first life-sized reconstruction of this dinosaur, was donated to the park.[23][87][88]
Дилофозавр was proposed as the state dinosaur of Arizona by a 9 year-old boy in 1998, but lawmakers suggested Сонорасавр instead, arguing that Дилофозавр was not unique to Arizona. A compromise was suggested that would recognize both dinosaurs, but the bill died when it was revealed that the Дилофозавр fossils had been taken without permission from the Navajo Reservation, and because they did not reside in Arizona anymore. Navajo Nation officials subsequently discussed how to get the fossils returned.[89][90] According to Mayor, one Navajo stated that they do not ask to get the fossils back anymore, but wondered why casts had not been made so the bones could be left, as it would be better to keep them in the ground, and a museum built so people could come to see them there.[86] 11 жастағы бала тағы да ұсыныс жасады Сонорасавр Аризонаның мемлекеттік динозавры ретінде 2018 ж.[90]
Әдебиеттер тізімі
- ^ «Dilophosaurus». Оксфорд сөздіктері Ұлыбритания сөздігі. Оксфорд университетінің баспасы. Алынған 21 қаңтар, 2016.
- ^ а б c г. e f ж сағ мен j к л м n o б q р с т сен v w х ж з аа Уэллс, С.П. (1984). "Dilophosaurus wetherilli (Динозаврия, Теропода), остеология және салыстыру ». Palaeontographica Abteilung A. 185: 85–180.
- ^ а б c Уэллс, С.П. (1954). «Аризонаның Кайента формациясындағы жаңа юра динозавры». Американың геологиялық қоғамының хабаршысы. 65 (6): 591–598. Бибкод:1954GSAB ... 65..591W. дои:10.1130 / 0016-7606 (1954) 65 [591: NJDFTK] 2.0.CO; 2.
- ^ Уэллс, С.П.; Гуралник, Р.П. (1994). "Дилофозавр ашылды «. ucmp.berkeley.edu. Калифорния университеті, Беркли. Мұрағатталды түпнұсқадан 2017 жылғы 8 қарашада. Алынған 13 ақпан, 2018.
- ^ а б c г. e f Найш, Д. (2009). Ұлы динозавр ашылымдары. Лондон, Ұлыбритания: A & C Black Publishers Ltd., 94–95 б. ISBN 978-1-4081-1906-8.
- ^ а б c г. e f ж сағ мен j к л м n Марш, А.д .; Роу, Т.Б. (2020). «Анатомиялық-филогенетикалық кешенді бағалау Dilophosaurus wetherilli (Динозаврия, Теропода) Аризонаның солтүстігіндегі Кайента формациясындағы жаңа үлгілердің сипаттамасымен «. Палеонтология журналы. 94 (S78): 1–103. дои:10.1017 / jpa.2020.14. S2CID 220601744.
- ^ а б c г. e f Уэллс, С.П. (1970). "Дилофозавр (Рептилия: Саурисчия), динозаврдың жаңа атауы ». Палеонтология журналы. 44 (5): 989. JSTOR 1302738.
- ^ а б Уэллс, С.П.; Гуралник, Р.П. (1994). «Дилофозавр туралы мәліметтер». ucmp.berkeley.edu. Калифорния университеті, Беркли. Мұрағатталды түпнұсқадан 2017 жылғы 2 тамызда. Алынған 13 ақпан, 2018.
- ^ а б Психойос, Л .; Кнеббер, Дж. (1994). Динозаврларды аулау. Лондон, Ұлыбритания: Касселл. 86–89 бет. ISBN 978-0679431244.
- ^ а б Раухут, О.В. (2004). «Базальды теропод динозаврларының өзара байланысы және эволюциясы». Палеонтологиядағы арнайы құжаттар. 69: 213.
- ^ а б Глут, Д.Ф. (1997). Динозаврлар: Энциклопедия. Джефферсон: McFarland & Company, Inc. 347–350 бб. ISBN 978-0786472222.
- ^ а б c г. Гей, Р. (2001). Жаңа үлгілері Dilophosaurus wetherilli (Динозавр: Теропода) Аризонаның солтүстігіндегі юра дәуірінің Кайента қалыптасуынан. Батыс омыртқалы палеонтологтар қауымдастығы жыл сайынғы кездесу. 1. Меса, Аризона. б. 1.
- ^ а б c г. Каррано, М.Т .; Бенсон, Р.Б.Ж .; Сампсон, С.Д. (2012). «Тетанураның филогениясы (Динозаврия: Теропода)». Систематикалық палеонтология журналы. 10 (2): 211–300. дои:10.1080/14772019.2011.630927. S2CID 85354215.
- ^ а б c Тыкоски, Р.С. (2005). Целофифоидты тероподтардың анатомиясы, онтогенезі және филогениясы (Тезис). Техас университеті. 1–232 бет - UT кітапханалары арқылы: Электрондық тезистер мен диссертациялар.
- ^ Пикрелл, Дж. (7 шілде, 2020). "Юра паркі осы көрнекті динозавр туралы барлық дерлік қате кетті ». ұлттық географиялық. Ғылым. Алынған 12 шілде, 2020.
- ^ а б Гей, Р. (2005). «Ерте юра тероподты динозаврдағы жыныстық диморфизмге дәлел, Дилофозавр және басқа байланысты формалармен салыстыру ». Карпентерде К. (ред.). Жыртқыш динозаврлар. Блумингтон, IN: Индиана Университетінің баспасы. 277–283 беттер. ISBN 978-0-253-34539-4.
- ^ Mortimer, M. (мамыр 2010). «Пикерингтің таксоны 6: Дилофозавр селекция «. Theropoddatabase.blogspot.com. Мұрағатталды түпнұсқасынан 2017 жылғы 29 желтоқсанда. Алынған 29 желтоқсан, 2017.
- ^ Хендриккс, С .; Матеус О .; Эванс, Алистер Роберт (2014). "Torvosaurus gurneyi n. sp., Еуропадағы ең үлкен құрлықтағы жыртқыш және құс емес тероподтардағы максиларлы анатомияның ұсынылған терминологиясы ». PLOS ONE. 9 (3): e88905. Бибкод:2014PLoSO ... 988905H. дои:10.1371 / journal.pone.0088905. PMC 3943790. PMID 24598585.
- ^ Ху, С. (1993). «Пайда болуы туралы қысқаша есеп Дилофозавр Юньнань провинциясының Цзинь округінен ». Vertebrata PalAsiatica. 1 (қытай тілінде) (1 ред.) 31: 65–69.
- ^ а б Ламанна, МС .; Хольц, ТР, кіші; Додсон, П. (1998). «Қытай тероподты динозаврды қайта бағалау Dilophosaurus sinensis". Омыртқалы палеонтология журналы. Мақалалардың тезистері, елу сегізінші жылдық кездесу, омыртқалы палеонтология қоғамы. 18 (3): 57–58. JSTOR 4523942.
- ^ Син, Л .; Белл, П.Р .; Ротшильд, Б.М .; Ран, Х .; Чжан, Дж .; Дон, З .; Чжан, В .; Currie, PJ (2013). «Тістің жоғалуы және альвеолярлы қайта құру Sinosaurus triassicus (Динозаврия: Теропода) Люфен бассейнінің төменгі юра қабаттарынан, Қытай ». Қытай ғылыми бюллетені. 58 (16): 1931. Бибкод:2013ChSBu..58.1931X. дои:10.1007 / s11434-013-5765-7.
- ^ Ван, Гуо-Фу; Сіз, Хай-Лу; Пан, Ши-Ганг; Ванг, Дао (2017). «Қытайдағы Юннань провинциясының ерте юра дәуірінен шыққан жаңа крест тәрізді динозавр». Vertebrata PalAsiatica. 55 (2): 177–186.
- ^ а б c г. e f ж Пол, Г.С. (1988). Әлемнің жыртқыш динозаврлары. Нью-Йорк, Нью-Йорк: Саймон және Шустер. бет.258, 267–271. ISBN 978-0-671-61946-6.
- ^ Holtz, TR, Jr (2012). Динозаврлар: барлық жастағы динозаврларды сүйетіндерге арналған ең толық, заманауи энциклопедия. Нью-Йорк, Нью-Йорк: Кездейсоқ үй. б.81. ISBN 978-0-375-82419-7.
- ^ Пол, Г.С. (2010). Динозаврларға арналған Принстондағы далалық нұсқаулық. Принстон, NJ: Принстон университетінің баспасы. б.75. ISBN 978-0-691-13720-9.
- ^ а б Джерлиńски, Г. (1996). «Массачусетстің төменгі Юра дәуірінен қалған тероподтағы қауырсын тәрізді әсер». Солтүстік Аризона бюллетенінің мұражайы. 60: 179–184.
- ^ а б Кундрат, М. (2004). «Тероподтар қашан қауырсынды болды? - Алдын ала айғақтарАрхеоптерикс қауырсынды қосымшалар »деп аталады. Тәжірибелік зоология журналы В бөлімі: Молекулалық және даму эволюциясы. 302B (4): 355–364. дои:10.1002 / jez.b.20014. PMID 15287100.
- ^ а б Локли, М .; Мацукава М .; Цзянцзюнь, Л. (2003). «Тероподтарды таксономиялық джунгилерде круиздеу: метатарсальды және ишиальды әсерлері бар іздерді икнологиялық және ичнотаксономикалық зерттеу». Ичнос. 10 (2–4): 169–177. дои:10.1080/10420940390256249. S2CID 128759174.
- ^ а б Мартин, А.Ж .; Rainforth, EC (2004). «Тероподтың демалатын ізі, сонымен қатар локомотив ізі: Хичкоктың AC 1/7 үлгісін зерттеу». Американың геологиялық қоғамы. Бағдарламалармен рефераттар. 36 (2): 96. мұрағатталған түпнұсқа 2004 жылғы 31 мамырда.
- ^ а б c г. e f ж Тыкоски, Р.С .; Роу, Т. (2004). «Цератозаврия». Вейшампельде Д.Б .; Додсон, П .; Osmolska, H. (ред.). Динозавр (2 басылым). Беркли, Калифорния: Калифорния университетінің баспасы. 47–70 бет. ISBN 978-0-520-24209-8.
- ^ Уэллс, С.П. (1983). «Терропод астрагаласындағы сүйектенудің екі орталығы». Палеонтология журналы. 57 (2): 401. JSTOR 1304663.
- ^ Уэллс, С.П.; Ұзын, Р.А. (1974). «Терропод динозаврларының тарсусы» [Оңтүстік Африка мұражайының жылнамалары]. Annale van die Suid-Afrikaanse мұражайы. 64: 191–218. ISSN 0303-2515.
- ^ Хольц, ТР, кіші (1994). «Тираннозаврдың филогенетикалық жағдайы: тероподтардың систематикасына әсері». Палеонтология журналы. 68 (5): 1100–1117. дои:10.1017 / S0022336000026706. JSTOR 1306180.
- ^ Мадсен, Дж. Х .; Уэллс, С.П. (2000). «Ceratosaurus (Dinosauria, Theropoda): қайта қаралған остеология». Юта геологиялық қызметі: 1–89. 41293.
- ^ а б Йейтс, А.М. (2005). «Оңтүстік Африканың ерте юра дәуірінен шыққан жаңа теропод динозавры және оның тероподтардың ерте эволюциясы үшін салдары». Палеонтология Африка. 41: 105–122. ISSN 0078-8554.
- ^ Смит, Н.Д.; Маковики, П.Ж .; Хаммер, В.Р .; Currie, PJ (2007). «Остеология Cryolophosaurus ellioti (Динозаврия: Теропода) Антарктиданың ерте Юра дәуірінен және теропод эволюциясының салдары ». Линне қоғамының зоологиялық журналы. 151 (2): 377–421. дои:10.1111 / j.1096-3642.2007.00325.x.
- ^ Хендриккс, С .; Хартман, С.А .; Mateus, O. (2015). «Құс емес тероподтардың ашылуларына шолу және классификация». PalArch журналы омыртқалы палеонтология. 12 (1): 73.
- ^ Захнер, М .; Brinkmann, W. (2019). «Швейцариядан келген триастық авеространды теропод және динозаврлардың алғашқы эволюциясы». Табиғат экологиясы және эволюциясы. 3 (8): 1146–1152. дои:10.1038 / s41559-019-0941-з. PMC 6669044. PMID 31285577.
- ^ а б Уэллс, С.П. (1971). «Аризонаның солтүстігіндегі Кайента формациясынан алынған динозавр іздері». Үстірт. 44: 27–38.
- ^ Моралес, М .; Булкли, С. (1996). «Теропод динозаврына қарағанда палеоихнологиялық дәлелдемелер Дилофозавр Төменгі Юраның Кайента түзілуінде. Континентальды юра ». Солтүстік Аризона бюллетенінің мұражайы. 60: 143–145.
- ^ Джерлиńски, Г. (1991). «Қасиетті Крест Тауларының Ерте Юрасынан алынған жаңа динозавр ичнотакса». Палеогеография, палеоклиматология, палеоэкология. 85 (1–2): 137–148. Бибкод:1991PPP .... 85..137G. дои:10.1016 / 0031-0182 (91) 90030-U.
- ^ Джерлини, Г .; Ahlberg, A. (1994). «Швецияның оңтүстігіндегі Хоганас формациясындағы кеш триас және ерте юрасалық динозавр іздері». Ичнос. 3 (2): 99. дои:10.1080/10420949409386377.
- ^ Weems, R.E. (2003). "Платеозавр аяқ құрылымы үшін жалғыз трекмейкер ұсынады Eubrontes және Гигандип іздері ». Ле-Турно, П.М .; Олсен, П.Е. (ред.). Шығыс Солтүстік Америкадағы Пангеяның үлкен рифт аңғарлары. 2. Нью-Йорк: Колумбия университетінің баспасы. бет.293 –313. ISBN 978-0231126762.
- ^ Rainforth, EC (2003). «Ерте юра динозаврының ішногенін қайта қарау және қайта бағалау Отозум". Палеонтология. 46 (4): 803–838. дои:10.1111/1475-4983.00320.
- ^ а б Weems, R.E. (2006). «Қолжазба басылымы Кәмелетке толмаған Kayentapus; оның ерте мезозойлық сауришиялық динозаврлардың биомеханикасы мен ичнотаксономиясына әсері ». Нью-Мексико Жаратылыстану тарихы және ғылыми мұражайы. 37: 369–378.
- ^ Лукас, С.Г .; Клейн, Х .; Локли, МГ .; Шпилманн, Дж .; Джерлинский, Г.Д .; Хант, А.П .; Таннер, Л.Х. (2006). «Терноподтық изиннің триас-юра стратиграфиялық таралуы Eubrontes". Нью-Мексико Жаратылыстану тарихы және ғылыми мұражайы. 37. 265.
- ^ а б c Милнер, Эндрю РК .; Харрис, Дж .; Локли, МГ .; Кирклэнд, Дж .; Мэтьюз, Н.А .; Harpending, H. (2009). «Ерте юра тероподты динозаврдың тынығу ізімен анықталған құс тәрізді анатомиясы, қалпы және мінез-құлқы». PLOS ONE. 4 (3): e4591. Бибкод:2009PLoSO ... 4.4591M. дои:10.1371 / journal.pone.0004591. PMC 2645690. PMID 19259260.
- ^ Weems, R.E. (2019). «Екі аяқты прозауроподтарға дәлел Eubrontes трек жасаушылар ». Ичнос. 26 (3): 187–215. дои:10.1080/10420940.2018.1532902. S2CID 133770251.
- ^ Баккер, Р.Т. (1986). Динозаврлардың бидғаттары. Нью-Йорк, Нью-Йорк: Уильям Морроу. бет.263 –264. ISBN 978-0-8217-5608-9.
- ^ Терриен, Ф .; Хендерсон, Д .; Ruff, C. (2005). «Мені тісте - тероподтардың төменгі жақ сүйектерінің биомеханикалық модельдері және тамақтану тәртібіне салдары». Карпентерде К. (ред.) Жыртқыш динозаврлар. Индиана университетінің баспасы. 179–230 бб. ISBN 978-0-253-34539-4.
- ^ Милнер, А .; Kirkland, J. (2007). «Джонсон фермасындағы Георгий динозаврларын табу алаңында динозаврларды аулауға арналған іс» (PDF). Юта штатындағы геологиялық зерттеудің жазбалары. 39: 1–3.
- ^ Марш, А.д .; Роу, Т.Б. (2018). «Сауроподоморфтың анатомиясы және систематикасы Sarahsaurus aurifontanalis Ерте Юра Кайента формациясынан ». PLOS ONE. 13 (10): e0204007. Бибкод:2018PLoSO..1304007M. дои:10.1371 / journal.pone.0204007. PMC 6179219. PMID 30304035.
- ^ а б Черкас, С.Ж .; Czerkas, SA (1990). Динозаврлар: ғаламдық көрініс. Лимпсфилд: Айдаһар әлемі. б. 208. ISBN 978-0-7924-5606-3.
- ^ Пол, Г.С., басылым. (2000). «Түс бөлімі». Американдық ғылыми динозаврлар кітабы. Нью-Йорк, Нью-Йорк: Сент-Мартин баспасөзі. б. 216. ISBN 978-0-312-31008-0.
- ^ Senter, P .; Робинс, Дж. (2005). «Теропод динозаврының алдыңғы аясындағы қозғалыс ауқымы Acrocanthosaurus atokensis, және жыртқыш мінез-құлықтың салдары ». Зоология журналы. 266 (3): 307–318. дои:10.1017 / S0952836905006989.
- ^ Senter, P .; Робинс, Дж. (2015). «Динозаврлардың скапула мен алдыңғы аяқтарының тынығу бағыттары: қайта құру мен мұражай монтаждарына әсер ететін сандық талдау». PLOS ONE. 10 (12): e0144036. Бибкод:2015PLoSO..1044036S. дои:10.1371 / journal.pone.0144036. PMC 4684415. PMID 26675035.
- ^ а б c г. Senter, P .; Салливан, C. (2019). «Теропод динозаврының алдыңғы аяқтары Dilophosaurus wetherilli: Қозғалыс ауқымы, палеопатология мен жұмсақ тіндердің әсері және дистальды карпальды сүйектің сипаттамасы ». Palaeontologia Electronica. дои:10.26879/900.
- ^ Кумбс, В.П. (1990). «Динозаврлардың мінез-құлық үлгілері». Вейшампельде Д.Б .; Осмольска, Х .; Додсон, П. (ред.) Динозавр (1-ші басылым). Беркли, Калифорния: Калифорния университетінің баспасы. б. 42. ISBN 978-0-520-06727-1.
- ^ Падиан, К .; Хорнер, Дж.Р. (2011). «Динозаврлардағы» таңқаларлық құрылымдардың «эволюциясы: биомеханика, жыныстық сұрыптау, әлеуметтік сұрыптау немесе түрлерді тану?». Зоология журналы. 283 (1): 3–17. дои:10.1111 / j.1469-7998.2010.00719.x.
- ^ Кнелл, Р.Дж .; Sampson, S. (2011). «Динозаврлардағы таңқаларлық құрылымдар: түрлерді тану немесе жыныстық сұрыптау? Падиан мен Хорнерге жауап» (PDF). Зоология журналы. 283 (1): 18–22. дои:10.1111 / j.1469-7998.2010.00758.x.
- ^ Hone, D.W.E .; Найш, Д. (2013). «» Түрлерді тану гипотезасы «авиалан емес динозаврлардағы экстрагирленген құрылымдардың болуы мен эволюциясын түсіндірмейді». Зоология журналы. 290 (3): 172–180. дои:10.1111 / jzo.12035.
- ^ Ткач, Дж.С. (19 қазан 1996). Көп элементтерді остеохистологиялық талдау Dilphosaurus wetherilli. Елу алтыншы жылдық кездесу, Омыртқалы палеонтология қоғамы. Омыртқалы палеонтология журналы. Қысқаша тезистер. 16 (3). Американдық табиғи тарих мұражайы, Нью-Йорк, Нью-Йорк. 1А – 80А беттер. JSTOR 4523755.
- ^ Молнар, Р.Е. (2001). «Теропод палеопатологиясы: әдеби зерттеу». Танкеде, Д.Х .; Ағаш ұстасы, К. (ред.). Мезозой омыртқалы тіршілігі. Блумингтон, IN: Индиана Университетінің баспасы. бет.337 –363. ISBN 978-0-253-33907-2.
- ^ Ротшильд, Б .; Танке, Д.Х .; Форд, Т.Л. (2001). «Тероподтық стресстің сынуы және сіңірдің авульсиясы белсенділіктің белгісі ретінде». Танкеде, Д.Х .; Ағаш ұстасы, К. (ред.). Мезозой омыртқалы тіршілігі. Блумингтон, IN: Индиана Университетінің баспасы. бет.331 –336. ISBN 978-0253339072.
- ^ а б Senter, P .; Хуенгст, С.Л .; Heymann, D. (2016). «Рекордтық ауырсыну: тероподты динозаврдағы алдыңғы сүйек ауруының ең көп саны және әртүрлілігі». PLOS ONE. 11 (2): e0149140. Бибкод:2016PLoSO..1149140S. дои:10.1371 / journal.pone.0149140. PMC 4765892. PMID 26909701.
- ^ а б Дж. М., Кларк; Фастовский, Д.Е. (1986). «Аризонаның солтүстігіндегі Глен Каньон тобының омыртқалы биостратиграфиясы». Фрейзерде, Н.С .; Сьюс, Х.Д. (ред.). Динозаврлар дәуірінің басталуы: Триас-Юра шекарасындағы фауналық өзгеріс. Кембридж, Ұлыбритания: Кембридж университетінің баспасы. 285–301 бет. ISBN 978-0-521-36779-0.
- ^ а б Харшбаргер, Дж. В .; Репенинг, C. А .; Ирвин, Дж. Х. (1957). «Навахо елінің ең жоғарғы триас және юра жыныстарының стратиграфиясы». Нью-Мексико геологиялық қоғамы. 291: 98–114.
- ^ а б c Лукас, С.Г .; Хеккерт, А.Б .; Таннер, Л.Х. (2005). «Аризонаның Юра дәуіріндегі қазба омыртқалылар және Глен Каньон тобының жасы». Нью-Мексико Табиғат тарихы мұражайы бюллетендері. 29: 95–104.
- ^ а б c Луттрелл, П.Р .; Моралес, М. (1993). «Moenkopi Wash арасындағы алшақтықты жою: литостратиграфиялық корреляция». Солтүстік Аризона бюллетенінің мұражайы. 59: 111–127.
- ^ а б Дженкинс, Ф.А .; Шубин, Н.Х. (1998). «Просалирус биті және ануран-каудопельвик механизмі ». Омыртқалы палеонтология журналы. 18 (3): 495–510. дои:10.1080/02724634.1998.10011077.
- ^ Ригби, Дж .; Гамблин, ВК .; Матени, Р .; Уэльс, S.L. (1971). «Колорадо өзеніне нұсқаулық: 3-бөлім, Моабтан Хайтке, Юта, Каньонлэнд ұлттық паркі арқылы» (PDF). Бригам Янг Университетінің зерттеу жұмыстары, Геология сериясы. 18 (2): 7.
- ^ Лукас, С.Г .; Таннер, Л.Х. (2007). «АҚШ-тың оңтүстік Колорадо үстіртіндегі триас-юра кезеңінің тетрапод биостратиграфиясы және биохронологиясы». Палеогеография, палеоклиматология, палеоэкология. 244 (1–4): 242–256. Бибкод:2007PPP ... 244..242L. дои:10.1016 / j.palaeo.2006.06.030.
- ^ Кертис, К .; Падиан, К. (1999). «Аризонаның солтүстік-шығысындағы Кайента түзілуінен ерте юра дәуіріндегі микро омыртқалылар фаунасы: триас-юра шекарасы арқылы микрофуналдың өзгеруі». PaleoBios. 19 (2): 19–37.
- ^ а б Дженкинс, Ф. А .; Кромптон, А .; Даунс, В.Р. (1983). «Аризонадан мезозойлық сүтқоректілер: сүтқоректілер эволюциясының жаңа дәлелі». Ғылым. 222 (4629): 1233–1235. Бибкод:1983Sci ... 222.1233J. дои:10.1126 / ғылым.222.4629.1233. PMID 17806725. S2CID 35607107.
- ^ Гей, Р. (2010). "Kayentavenator elysiae, Аризонаның ерте Юра дәуірінен алынған жаңа тетануран » Гей, Р. (ред.). Ерте мезозойлық тероподтар туралы ескертпелер. Lulu Press. 27-43 бет. ISBN 978-0557466160.
- ^ Роу, Т.Б .; Сьюс, Х.-Д .; Reisz, RR (2010). «Жаңа таксонның сипаттамасымен алғашқы солтүстікамерикалық сауроподоморфтық динозаврлардың таралуы және әртүрлілігі». Корольдік қоғамның еңбектері B: Биологиялық ғылымдар. 278 (1708): 1044–1053. дои:10.1098 / rspb.2010.1867. PMC 3049036. PMID 20926438.
- ^ Гамблин, А.Х .; Фостер, Дж.Р. (2000). «Үлкен баспалдақтағы ежелгі жануарлардың іздері мен іздері - Юта штатының оңтүстігінде Эскаланте ұлттық ескерткіші». Спринкелде, Д.А .; Чидси, ТК, кіші; Андерсон, П.Б. (ред.). Юта парктері мен ескерткіштерінің геологиясы. Солт-Лейк-Сити: Юта геологиялық қауымдастығы. 557–568 беттер. ISBN 978-1882054107.
- ^ Рейсдорф, А.Г .; Wuttke, M. (2012). «Модиенің қазба омыртқалылардағы опистотоникалық-қалыптық гипотезасын қайта бағалау, І бөлім: Рептилиялар - екі аяқты динозаврлардың тапономиясы Compsognathus longipes және Juravenator starki Солнхофен архипелагынан (Юра, Германия) ». Палеобиоалуантүрлілік және палеоорталар. 92 (1): 119–168. дои:10.1007 / s12549-011-0068-ж. S2CID 129785393.
- ^ Crichton, M. (1990). Юра паркі. Лондон: Random Century Group. бет.76–78. ISBN 978-0-394-58816-2.
- ^ Шей, Д .; Дункан, Дж. (1993). Юра саябағын құру. Нью-Йорк: Boxtree Ltd. 24, 35–36, 113 беттер. ISBN 978-1-85283-774-7.
- ^ Дункан, Дж. (2007). Уинстон эффектісі: Стэн Уинстон студиясының өнері мен тарихы. Лондон, Ұлыбритания: Титан кітаптары. 177–178 бб. ISBN 978-1-84576-150-9.
- ^ Bennington, JB (1996). «Фильмдегі қателіктер Юра паркі". Американдық палеонтолог. 4 (2): 4-7. Архивтелген түпнұсқа 2018 жылғы 2 қазанда. Алынған 28 желтоқсан, 2017.
- ^ Додсон, П. (1997). «Өлгендерді тірілту». Ғылым. 277 (5326): 644–645. дои:10.1126 / ғылым.277.5326.644b. JSTOR 2893254. S2CID 178195275.
- ^ Баккер, Р.Т. (2014). «Екі компас туралы ертегі: Не Юра паркі дино-анатомия туралы дұрыс және қате ». blog.hmns.org. Хьюстон жаратылыстану мұражайы. Алынған 18 тамыз, 2020.
- ^ Уэллс, С.П.; Гуралник, Р.П. (1994). "Дилофозавр, актер ». ucmp.berkeley.edu. Калифорния университеті, Беркли. Мұрағатталды түпнұсқадан 2018 жылғы 4 қаңтарда. Алынған 13 ақпан, 2018.
- ^ а б c г. Әкім, А. (2005). «Оңтүстік-батыс: қазба фетиштері мен монстрлерді өлтірушілер». Бірінші американдықтардың қазба аңыздары. Принстон, NJ: Принстон университетінің баспасы. 130–143 бет. ISBN 978-0691130491. JSTOR j.ctt4cgcs9.10.
- ^ Stone, M. (2017). «Коннектикут өзінің жаңа мемлекеттік динозаврын қарсы алады». CTBoom.com. Алынған 13 ақпан, 2018.
- ^ Роджерс, О. (2016). «Табылған динозавр жолдары магистральға қайта бағыт алып, мемлекеттік паркке апарады». Коннектикут тарихы. Алынған 3 қаңтар, 2018.
- ^ Moeser, C. (26 сәуір, 1998). «Аризонадағы динозаврлар туралы пікірталас нағыз Godzilla-ға айналды». Аризона Республикасы. Deseret News. Алынған 12 мамыр, 2020.
- ^ а б Геберс, С. (19.02.2018). «Аризонада ресми мемлекеттік динозавр жоқ, сондықтан 11 жасар жасөспірім - оны ұсынады Сонорасавр". Республика. Алынған 20 ақпан, 2018 - AZ Central арқылы.
Сыртқы сілтемелер
- «Дилофозавр!». Баяндамалы көрме «экскурсия». баяндаған Уэллс.CS1 maint: басқалары (сілтеме)













