Камарасавр - Camarasaurus
| Камарасавр | |
|---|---|
 | |
| Қаңқаға құйылған Юралық астурия мұражайы | |
| Ғылыми классификация | |
| Корольдігі: | Анималия |
| Филум: | Chordata |
| Клайд: | Динозаврия |
| Клайд: | Сауришия |
| Қосымша тапсырыс: | †Сауроподоморфа |
| Клайд: | †Сауропода |
| Клайд: | †Камарасауроморфа |
| Клайд: | †Camarasauridae |
| Субфамилия: | †Камарасауриндер Cope, 1878 жыл |
| Тұқым: | †Камарасавр Cope, 1877a |
| Түр түрлері | |
| †Camarasaurus supremus Cope, 1877a | |
| Түрлер | |
| |
| Синонимдер | |
| |
Камарасавр (/ˌкæмерəˈсɔːрəс/ KAM-әр-ə-SAWR-әс ) болды түр туралы төртбұрышты, шөпқоректі динозаврлар. Бұл алыптың ең кең тарағаны болды сауроподтар табуға болады Солтүстік Америка. Оның қалдықтары табылды Моррисонның қалыптасуы туралы Колорадо және Юта, .мен танысу Кейінгі юра дәуір (Киммеридгиан дейін Титониан кезеңдер), бұдан 155 - 145 миллион жыл бұрын.
Камарасавр доғал тұмсық пен доға тәрізді бас сүйектің ерекше төртбұрышты профилін ұсынды. Бұл табындарда, немесе, ең болмағанда, отбасылық топтарда жүрсе керек.
Атау «камералық кесіртке» дегенді білдіреді, ондағы қуыс камераларға сілтеме жасайды омыртқалар (Грек καμαρα (камара) «күмбезделген камера» немесе доға тәрізді қақпағы бар кез келген мағынаны білдіреді және σαυρος (саурос) «кесіртке» мағынасын білдіреді).
Сипаттама
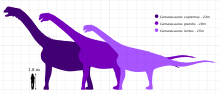
Камарасавр ең көп таралған және жиі сақталған сауопод динозаврларының бірі болып табылады. Ең көп таралған түрлердің максималды мөлшері, C. lentus, ұзындығы шамамен 15 м (49 фут) болды. Ең үлкен түрлері, C. supremus, максималды ұзындығы 23 м (75 фут), ал максималды есептік салмағы 47-ге жеттітонна (51.8 тоннаға жетеді ).[1][2]
Доғалы бас сүйегі туралы Камарасавр төртбұрышты және тұмсық көп болды fenestrae Палеонтологтар оны берік және жиі қалпына келтіреді. Ұзындығы 19 см (7,5 дюйм) тістер тәрізді болды қашау (спатуляция) және жақ бойымен біркелкі орналасады. Тістердің беріктігі оны көрсетеді Камарасавр жіңішке тістен гөрі ірі өсімдік материалын жеген шығар диплодоцидтер.[дәйексөз қажет ]
Әрбір алдыңғы аяқ бес саусақты, ішкі саусақ үлкен, үшкір болатын тырнақ. Көптеген сауоподтар сияқты алдыңғы аяқтары артқы аяқтарына қарағанда қысқа, бірақ жоғары орналасуы иық артқы жағында кішкене көлбеу болғанын білдірді.
Салмақты үнемдеу мақсатына қызмет ету, басқа сауоподтарда көрсетілгендей, көптеген омыртқалар ойық немесе «пневматикалық» болды; яғни, омыртқалар өкпеге байланысты ауа қапшықтарының күрделі жүйесі үшін өткелдер мен қуыстармен қаныққан. Бұл ерекшелік сол кезде аз түсінікті болатын Камарасавр ашылды, бірақ оның құрылымы «камералық кесіртке» дегенді білдіретін тіршілік иесінің атына шабыт болды. Мойын мен тепе-теңдік құйрығы осы мөлшердегі савопод үшін әдеттегіден қысқа болды. Камарасавр, кейбір басқа сауоподтар сияқты, ұлғайған жұлын жамбастың жанында. Палеонтологтар[ДДСҰ? ] бастапқыда бұл секунд деп санады ми, мүмкін, осындай үлкен жаратылысты үйлестіру үшін қажет. Шынында да, бұл қарқынды жүйке жүйесінің, мүмкін рефлекторлы немесе автоматты түрде әрекет етуі мүмкін болғанымен, бұл ми емес еді; мұндай ұлғаю омыртқалы жануарларда белгілі дәрежеде жиі кездеседі.[дәйексөз қажет ]
Камарасавр грандис басқа радиоприемникке қарағанда сауроподқа қарағанда күшті радиусы болды Вененозавр.[3]
Үлгісі Камарасавр SMA 0002 деп аталады (ол да тағайындалған) Катетозавр) Вайомингтің «Э.Т.» деп аталатын Хоу-Стефенс карьерінен жұмсақ тіндердің дәлелдері көрсетілген. Иек сызығының бойында жануарлардың қызылиегі болып көрінетін сүйектенген қалдықтар қалпына келтірілді, бұл оның терең тістердің қызыл иекпен жабылғандығын, тек тәждердің ұштары ғана шығып тұрғанын көрсетті. Тістер қайтыс болғаннан кейін, сағыздар тартылып, құрғап, ыдырау кезінде қатайған кезде ұяларынан итеріп шығарды. Үлгіні тексеру сонымен қатар тістердің қатты қабыршақпен жабылғанын және әр түрлі тұмсықпен жабылғанын көрсетеді, бірақ бұл белгілі емес.[4]
Ашылу тарихы


Алғашқы жазбасы Камарасавр бірнеше шашыраңқы болған кезде, 1877 жылдан келеді омыртқалар орналасқан Колорадо, Орамель В. Лукас. Оның ұзақ уақытқа созылған және келісімді бәсекелестігін іздеу (ретінде белгілі Сүйек соғысы ) бірге Отниель Чарльз Марш, палеонтолог Эдвард ішкіш сүйектерге ақы төлеп, тез қозғалып, сол жылы оларды атады. Өз кезегінде, Марш кейінірек кейбір сауоподты табыстарды атады Morosaurus grandis, бірақ қазіргі кезде палеонтологтардың көпшілігі оларды түрлер деп санайды Камарасавр Re: «Морозавр» agilis. Мұндай ат қою қақтығыстары екі қарсылас динозавр аңшыларының арасында жиі болды.
Толық қаңқасы Камарасавр сипатталмаған Чарльз В.Гилмор 1925 жылға дейін. Себебі бұл жас қаңқа КамарасаврАлайда, көптеген иллюстрациялар динозаврдың қазіргі кездегіден әлдеқайда аз екендігін көрсетеді.
Түрлер

The тип түрлері туралы Камарасавр Cope-тің түпнұсқа түрі, C. supremus («ең үлкен камералық кесіртке» дегенді білдіреді), 1877 жылы аталған. Табылғаннан кейінгі басқа түрлерге жатады C. grandis («үлкен камералы кесіртке») 1877 ж., C. lentus 1889 жылы және C. lewisi (бастапқыда жаңа тұқым ретінде сипатталған Катетозавр) 1988 ж.
C. supremus, оның аты айтып тұрғандай, белгілі ең ірі түрі болып табылады Камарасавр және соңғы юра дәуірінен белгілі ең массивті сауоподтардың бірі Моррисонның қалыптасуы. Үлкен көлемін қоспағанда, оны ажырату мүмкін болмады C. lentus. C. supremus тұтастай алғанда типке тән болмады және формацияның ең соңғы, жоғарғы бөліктерінен ғана белгілі. Екеуі де C. grandis және C. lentus аз болған, сонымен қатар Моррисонның алғашқы кезеңдерінде болған.
Стратиграфиялық дәлелдемелер хронологиялық дәйектіліктің үш түрдің физикалық айырмашылықтарына сәйкес келетіндігін көрсетеді және ол Моррисон формациясындағы эволюциялық прогрессияны сипаттайды. C. grandis ең ежелгі түрі және Моррисонның ең төменгі жыныстық қабаттарында болған. C. lentus бірге пайда болған кейінірек пайда болды C. grandis бірнеше миллион жыл бойы, мүмкін, екі түрдің жұлын анатомиясындағы айырмашылықтар ұсынған түрлі экологиялық қуыстарға байланысты. Кейінгі кезеңде C. grandis рок жазбадан жоғалып кетті, тек қалды C. lentus. Содан кейін C. lentusда жоғалып кетті; Сонымен қатар, C. supremus жоғарғы қабаттарында пайда болды. Түрлердің бұл бірден-бір сабақтастығы, сондай-ақ екеуінің бір-біріне өте жақын ұқсастығы оны көрсетеді C. supremus тікелей дамыған болуы мүмкін C. lentus, жануарлардың кейінірек тіршілік ететін популяциясын білдіреді.[5]
C. lewisi түрлері бастапқыда ретінде аталды Катетозавр левиси және кейінірек синониміне айналды Камарасавр. 2013 жылғы талдау екі тұқымды қайтадан бөлді.[6]
Жіктелуі

The ғылыми классификация туралы Камарасавр, пайдаланып Линней жүйесі, оң жақта орналасқан қорапта берілген, бірақ палеонтологтар арасында динозаврларды таксономиялық классификациялау әдісі ығыстырылып жатыр кладистика -шығарылған филогенетикалық таксономия.
D'Emic (2012) кейінгі Макронарияның оңайлатылған кладограммасы төменде көрсетілген:[7]
| Макронария |
| |||||||||||||||||||||||||||
Камарасавр болып саналады базальды макронарий, макронарийлердің бәрінен гөрі барлық макронарийлердің ортақ атасымен тығыз байланысты алынған сияқты формалар Брахиозавр.
Палеобиология

Отарлау
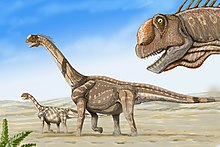
A қазба қалдықтары кейінгі юра дәуірінде, шамамен 150 миллион жыл бұрын (солтүстік-шығыста) бірге қайтыс болған екі ересек адам және ұзындығы 12,2 м (40 фут) жасөспірім бар. Вайоминг, АҚШ).[1 ескерту] Олардың денелерін аллювиалды балшықта соңғы тынығатын жеріне дейін (су тасқыны) өзен шайып кетеді деп болжанған. Сценарий осыны ұсынады Камарасавр саяхаттады табын немесе ең болмағанда отбасылық топтарда. Басқа учаскелерде, қазбалы камерасавр жұмыртқа ұқыпты орналастырылғаннан гөрі, сызықтардан табылды ұялар басқа динозаврлар сияқты, бұл көптеген савоподтар сияқты, Камарасавр оның жастарын өсірмеді.
Азықтандыру
Бұған дейін ғалымдар бұған кеңес берген Камарасавр және басқа сауроподтар жұтып қойған болуы мүмкін гастролиттер (тастар) асқазандағы тағамды ұнтақтауға көмектеседі, олар тым тегіс болған кезде оларды қалпына келтіреді немесе өткізеді. Асқазандағы тастардың пайда болғаны туралы жақында жүргізілген талдаулар бұлай болмағанын көрсетеді. Мықты, берік тістер Камарасавр көптеген сауроподтарға қарағанда дамыған және орташа 62 күнде ауыстырылатын (М. Д'Эмик және басқалар), бұл Камарасавр болуы мүмкін мастикалы жұтқанға дейін оның аузындағы тамақ белгілі бір дәрежеде.[8] Басқа табылулар осыны көрсетеді Камарасавр спп. басқа савроподтардан ерекшеленетін өсімдік жамылғысы, олардың бәсекелес болмай бір ортада болуына мүмкіндік берді.[9]
Өсу
Ұзын сүйек гистология зерттеушілерге нақты бір адамның жеткен жасын бағалауға мүмкіндік береді. Грибелер және басқалардың зерттеуі. (2013) ұзын сүйекті гистологиялық деректерді зерттеп, деген қорытындыға келді Камарасавр sp. CM 36664 салмағы 14247 килограмм (15,7 қысқа тонна), 20 жасында жыныстық жетілуге жетті және 26 жасында қайтыс болды.[10]
Метаболизм
Бүркіт т.б. орындалды изотоптық термометрия әр түрлі юралық савроподтардың тістерін жабатын эмальға, соның ішінде Камарасавр. 32.4-36.9 ° C температурасы алынды (90.3-98.4 ° F), бұл қазіргі сүтқоректілердің температурасымен салыстырылады.[11]
Палеопатология

A Камарасавр жамбас қалпына келді Динозавр ұлттық монументі Юта штатында гугинг жатады Аллозавр.[12]
1992 жылы жартылай C. grandis қаңқасы табылды Брайан Шағын Стегозавр карьері жақын Моррисон формациясы Канон-Сити, Колорадо.[13] Бұл үлгі DMNH 2908 ретінде каталогталған ішінара оң жақ иық сүйектерін және артқы және құйрық жағынан байланысты омыртқаларды сақтап қалды.[13] 2001 жылы Лори МакВинни, Кеннет Карпентер және Брюс Ротшильд иықта байқалған патологияның сипаттамасын жариялады.[14] Олар а жақстакортикальды зақымдану 25 - 18 см ені тоқылған талшықтарға ұқсайтын сүйектен жасалған.[15] Дегенмен тоқылған сүйек аксессуарлы тіс сүйегінде пайда болады, басқа жерлерде бұл жарақат немесе аурудың белгісі.[15] Тоқылған сүйектің «толқынды талшықты шумақтары» бағытына бағытталғандығы байқалды м. brachialis.[15] Зақымдану біріктіру және болмауы кеуектілік жақын және алыс шетінде периоститтің белсенді емделмегенін немесе емделгенін көрсетеді.[15] МакВинни және басқа зерттеушілер бұл жарақат жануар үшін үнемі қиындық тудыратын еді деп тұжырымдады.[16] Бұл бұлшықеттерге қысым жасар еді.[14] Бұл қысым бұлшық еттердің қан тамырлары мен нервтерін қысып, екі аяқтың да қимыл-қозғалысын төмендететін еді бүгу және экстензор бұлшықеттері.[14] Бұл әсер кедергі болар еді мм. brachialis, м. brachoradialis, және аз дәрежеде м. бицепс брахии иық сүйегіндегі зақымдану жағдайына дейін.[14] Зерттеушілер бұлшықеттің қабынуы және периостеум алдыңғы аяқтың төменгі аймағында да қосымша асқынулар тудырған болар еді.[17] Зақымдану ұзаққа созылуы мүмкін фасциит және миозистис.[14] Осы патологиялық процестердің жиынтық әсері аяқ-қолдың қозғалу қабілетіне орташа және ауыр әсер етеді және «тамақтану және жыртқыштардан қашу сияқты күнделікті әрекеттерді орындауды қиындатады».[16]
Патологияның себебін анықтауға көмектесу үшін Маквинни және басқа зерттеушілер а Томографиялық томография 3-мм қадаммен.[18] КТ-да массаның тұрақты екендігі анықталды радиодистенттілік және сүйектің қыртысынан а радиолентті түзу.[19] Стресс сынуы немесе остеомиелит немесе инфекциялық периостит сияқты инфекциялық процедуралар туралы ешқандай дәлел табылған жоқ.[18] Олар сондай-ақ жоққа шығарды остеохондрома өйткені иін осі иықтың вертикаль осіне қатысты 25 ° құрайды, ал остеохондрома иық өсіне қарай 90 ° -та пайда болған болар еді.[15] Сұйық зақымданудың шығу тегі үшін ғалымдар анықтаған басқа кандидаттар:
- Гипертрофиялық остеоартропатия - дегенмен, бұл шпор тәрізді процестің болуымен алынып тасталды[20]
- Остеоидты остеома - бірақ бұл демеуді түсіндірмейді остеобластикалық реакция[21]
- Жіңішке сынықтар немесе жіліншік стресс синдромы - ықтимал шығу тегі, өйткені көптеген белгілер ортақ болуы мүмкін, бірақ жіңішке буындар бұл септігін түсіндірмейді.[22]
- Myositis ossificans traumatica (Circscripta) - мүмкін, бірақ мүмкін емес көзі.[23]
- Авульсиялық жарақат - МакВинни және басқа зерттеушілер «бұлшық еттердің қайталанған шамадан тыс күштелуінен» туындаған авульсиялық жарақатты иық сүйегіндегі зақымданудың ең ықтимал көзі деп санады.[14] Зерттеушілер зақымдануды авульсиядан туындады деп санады м. brachialis «төмен қарай көлбеу эллипстік массаның» пайда болуын тудырады.[16] Сүйек сілкінісі негіздің жыртылуынан кейін остеобластикалық реакциядан туындады м. brachioradialis оның иілу қозғалысынан туындайды.[16]
Палеоэкология
Тіршілік ету ортасы

Моррисон формациясы, оның шығыс қапталында орналасқан Жартасты таулар, қазбаларға бай учаскенің отаны Кейінгі юра тау жынысы. Мұнда көптеген динозавр түрлерін, олардың туыстарын табуға болады Камарасавр сияқты Диплодокус, Апатозавр, және Брахиозавр, бірақ камарасаврлар динозаврлардың түзілуінде ең көп кездеседі.[24] Бірқатар толық қаңқалар алынды Колорадо, Нью-Мексико, Юта, және Вайоминг, әдетте 2-6 стратиграфиялық аймақтарда кездеседі.[25] Сәйкес радиометриялық танысу, Моррисон шөгінді қабаттары негізінен 156,3 миллион жыл бұрын (Мя), жоғарғы жағында 146,8 Мяға дейін созылады, бұл оны кейінге қалдырады Оксфордиан, Киммеридгиан және ерте Титониан кезеңдері кейінгі юра кезеңі.[26][27] Оның ортасы ретінде түсіндіріледі полимаридті айқын дымқыл және құрғақ маусымдар.
Динозавр және қазба қалдықтарын іздеу әсіресе Моррисон бассейнінде кездеседі, ол Нью-Мексикодан Альберта мен Саскачеванға дейін созылып, Рокки тауларының алдыңғы сілемдеріне дейінгі ізбасарлар батысқа қарай ығыса бастаған кезде пайда болды. Олардың шығысқа бағытталған эрозияланған материалы дренажды бассейндер ағындармен және өзендер және депонирленген батпақты ойпаттар, көлдер, өзен арналары және жайылмалар.[28] Жас түзілімі жасына қарай Solnhofen әктас Германиядағы қалыптасу және Тендагуру формациясы жылы Танзания. 1877 жылы ол орталыққа айналды Сүйек соғысы, алғашқы палеонтологтар арасындағы қазба жинақтайтын бәсекелестік Отниель Чарльз Марш және Эдвард ішкіш.
Палеофауна

Моррисон формациясы сияқты орасан зор сауропод динозаврлары басым болатын орта мен уақытты жазады Барозавр, Диплодокус, Апатозавр, Бронтозавр, және Брахиозавр. Қатар тұрған динозаврлар Камарасавр шөп қоректілерді қосады орнитисчилер Кемптозавр, Гаргойлеозавр, Dryosaurus, Стегозавр, және Отниелозавр. Бұл палео-ортадағы жыртқыштарға тероподтар Саурофаганакс, Торвозавр, Ceratosaurus, Маршосавр, Стокозозавр, Орнитолест,[29] және Аллозавр ол тероподтардың 75% -на дейін жетіп, жоғарғы жағында болды трофикалық деңгей Моррисон тамақтану торы.[30][31] Камарасавр әдетте сол сайттарда кездеседі Аллозавр, Апатозавр, Стегозавр, және Диплодокус.[32]
Осы аймақтың басқа ағзалары кіреді қосжапырақтылар, ұлы, сәулелі балықтар, бақалар, саламандрлар, тасбақалар, сфенодонттар, кесірткелер, құрлықтағы және судағы крокодиломорфтар, және бірнеше түрлері птерозаврлар сияқты Harpactognathus және Мезадактил. Ерте сүтқоректілер болған докодонттар (сияқты Docodon ), мультитуберкуляттар, симметродонттар және триконодонттар. Пайдалы қазбалар кезеңнің флорасын анықтады жасыл балдырлар, саңырауқұлақтар, мүктер, жылқылар, циклдар, гинкго, және бірнеше отбасы қылқан жапырақты ағаштар. Өсімдік жамылғысы өзен жағалауларындағы ормандардан әр түрлі болды папоротниктер, және папоротниктер (галерея ормандары ), папоротникке саванналар сияқты кездейсоқ ағаштармен Араукария - қылқан жапырақты ағаш тәрізді Brachyphyllum.[33]
Ескертулер
- ^ Омыртқалы палеонтология бөлімі қазған Канзас университетінің табиғи тарих мұражайы және биоалуантүрлілік орталығы, 1997 және 1998 жылдардағы далалық маусымдарда.
Пайдаланылған әдебиеттер
- ^ Бентон, Майкл Дж. (2012). Тарихқа дейінгі өмір. Эдинбург, Шотландия: Дорлинг Киндерсли. 270–271 бет. ISBN 978-0-7566-9910-9.
- ^ Фостер, Джон (2007). Юра Батысы: Моррисон формациясының динозаврлары және олардың әлемі. Индиана университетінің баспасы. 201, 248 б. ISBN 978-0-253-34870-8.
- ^ «Forelimb», Tidwell, Carpenter, and Meyer (2001). 148 бет.
- ^ «Камарасаврды аузына қарау».
- ^ "Camarasaurus grandis, «Фостер (2007). 204 бет.
- ^ Mateus, O., & Tschopp E. (2013). Катетозавр жарамды сауроподтар тұқымдасы және салыстыру Камарасавр. Омыртқалы палеонтология журналы, бағдарлама және тезистер, 2013. 173.
- ^ D'Emic, M. D. (2012). «Титанозавр формалы сауропод динозаврларының алғашқы эволюциясы» (PDF). Линне қоғамының зоологиялық журналы. 166 (3): 624–671. дои:10.1111 / j.1096-3642.2012.00853.x.
- ^ Wings and Sander (2006).
- ^ «Үлкен динозаврлардағы тістерді ауыстырудың жоғары жылдамдығы олардың эволюциялық жетістігіне ықпал етті».
- ^ Griebeler, EM; Клейн, Н; Sander, PM (2013). «Сауроподоморфтық динозаврлардың қартаюы, жетілуі және өсуі ұзын сүйектердің гистологиялық деректерін қолдану арқылы өсу қисықтарынан азайтылған: әдістемелік шектеулер мен шешімдерді бағалау». PLOS ONE. 8 (6): e67012. Бибкод:2013PLoSO ... 867012G. дои:10.1371 / journal.pone.0067012. PMC 3686781. PMID 23840575.
- ^ Орел, Р.А .; Түткен, Т .; Мартин, Т.С .; Трипати, А.К .; Фрикке, ХК; Коннели, М .; Цифелли, Р.Л .; Eiler, JM (2011-07-22). «Динозаврлардың дене температурасы изотоптық (13С-18О) органикалық қазбалардан алынған биоминералдардан анықталады». Ғылым. 333 (6041): 443–445. Бибкод:2011Sci ... 333..443E. дои:10.1126 / ғылым.1206196. PMID 21700837. S2CID 206534244.
- ^ «Камарасавр», Додсон және т.б. 56-бет.
- ^ а б «Кіріспе», МакВинни, Ұста және Ротшильд (2001); 365 бет.
- ^ а б c г. e f «Абстракт», МакВинни, Ұста және Ротшильд (2001); 364 бет.
- ^ а б c г. e «Сипаттама», МакВинни, Ұста және Ротшильд (2001); 367 бет.
- ^ а б c г. «Қорытындылар», МакВинни, Ұста және Ротшильд (2001); 376 бет.
- ^ «Сипаттама», МакВинни, Ұста және Ротшильд (2001); 369 бет.
- ^ а б «Сипаттама», МакВинни, Ұста және Ротшильд (2001); 370 бет.
- ^ «Сипаттама», МакВинни, Ұста және Ротшильд (2001); 370-371 беттер.
- ^ «Талқылау», МакВинни, Ұста және Ротшильд (2001); 373-373 беттер.
- ^ «Талқылау», МакВинни, Ұста және Ротшильд (2001); 373-374 беттер.
- ^ «Талқылау», МакВинни, Ұста және Ротшильд (2001); бет. 374.
- ^ «Талқылау», МакВинни, Ұста және Ротшильд (2001); 374-375 беттер.
- ^ "Camarasaurus supremus, «Фостер (2007). 201-бет.» Молшылықтар мен алуан түрлілік «, сонда. 248-бет.
- ^ «Қосымша», Фостер (2007). 328 бет.
- ^ Трухильо, К.С .; Чемберлен, К.Р .; Strickland, A. (2006). «Биостратиграфиялық корреляцияға байланысты оңтүстік-шығыс Вайомингтің Жоғарғы Юра Моррисонының қалыптасуына арналған SHRIMP талдауынан Oxfordian U / Pb қартаюы». Америка геологиялық қоғамы рефераттар бағдарламаларымен. 38 (6): 7.
- ^ Bilbey, SA (1998). «Кливленд-Ллойд динозавр карьері - жасы, стратиграфиясы және тұндыру ортасы». Карпентерде К .; Чуре Д .; Киркланд, Дж. (ред.). Моррисондық формация: пәнаралық зерттеу. Қазіргі геология. 22. Тейлор және Фрэнсис тобы. 87-120 бет. ISSN 0026-7775.
- ^ Рассел, Дейл А. (1989). Уақыттағы Одиссея: Солтүстік Американың динозаврлары. Минокуа, Висконсин: NorthWord Press. 64–70 бет. ISBN 978-1-55971-038-1.
- ^ Фостер, Дж. (2007). «Қосымша». Юра Батысы: Моррисон формациясының динозаврлары және олардың әлемі. Индиана университетінің баспасы. 327-329 бет.
- ^ Фостер, Джон (2007). «Allosaurus fragilis". Юра Батысы: Моррисон формациясының динозаврлары және олардың әлемі. Блумингтон, Индиана: Индиана университетінің баспасы. 170–176 бет. ISBN 978-0-253-34870-8. OCLC 77830875.
- ^ Фостер, Джон Р. (2003). Моррисон формациясындағы омыртқалы фаунаның палеоэкологиялық талдауы (жоғарғы юра), Рокки таулы аймағы, АҚШ. Нью-Мексико Табиғат тарихы және ғылыми мұражайы бюллетені, 23. Альбукерке, Нью-Мексико: Нью-Мексико Табиғи тарих және ғылым мұражайы. б. 29.
- ^ Додсон, Питер; Бехренсмейер, А.К .; Баккер, Роберт Т .; McIntosh, Джон С. (1980). «Юра Моррисон формациясындағы динозавр төсектерінің тапономиясы және палеоэкологиясы». Палеобиология. 6 (2): 208–232. дои:10.1017 / S0094837300025768.
- ^ Ағаш ұстасы, Кеннет (2006). «Үлкеннің ең үлкені: мега-савроподты сыни тұрғыдан қайта бағалау Amphicoelias fragillimus«. Фостерде Джон Р.; Лукас, Спенсер Г. (ред.). Палеонтология және жоғарғы юраның моррисон түзілуінің геологиясы. Нью-Мексико Жаратылыстану тарихы және ғылыми мұражайы. 36. Альбукерке, Нью-Мексико: Нью-Мексико Табиғи тарих және ғылым мұражайы. 131-138 бб.
Дереккөздер
- Cope, E. D. (1877a). «Колорадо штатындағы Дакота эопокынан алып сауриге». Палеонтологиялық бюллетень (25): 5–10.
- Cope, E. D. (1877b). «Колорадоның Дакота төсегінен шыққан рептилийлерде». Палеонтологиялық бюллетень (26): 193–197.
- Cope, E. D. (1877c). «Амфикоэлия туралы, Колорадо Дакота дәуіріндегі сауриялықтардың бір тұқымы». Американдық философиялық қоғамның еңбектері (17): 242–246.
- Cope, E. D. (1878). «Дакота Колорадо дәуірінің омыртқаларында». Палеонтологиялық бюллетень. 17 (28): 233–247. JSTOR 982303.
- Cope, E. D. (1879). «Жаңа юра динозавры». Американдық натуралист. 13: 401–404.
- Марш, О.С (1877). «Юра формациясынан шыққан жаңа динозаврлық рептилиялар туралы хабарлама». Американдық ғылым журналы. 3 серия. 14 (84): 514–516. Бибкод:1877AmJS ... 14..514M. дои:10.2475 / ajs.s3-14.84.514. S2CID 130488291.
- Марш, О.С (1878a). «Жаңа динозавр рептилиялары туралы хабарлама». Американдық ғылым журналы. 3 серия. 15 (87): 241–244. Бибкод:1878AmJS ... 15..241M. дои:10.2475 / ajs.s3-15.87.241. S2CID 131371457.
- Марш, О.С (1878б). «Американдық юра динозаврларының негізгі кейіпкерлері, I бөлім». Американдық ғылым журналы. 3 серия. 16 (95): 411–416. дои:10.2475 / ajs.s3-16.95.411. hdl:2027 / hvd.32044107172876. S2CID 219245525.
- Марш, О.С (1889). «Еуропа мен Американың динозаврларының негізгі формаларын салыстыру». Американдық ғылым журналы. 3 серия. 37 (220): 323–330. дои:10.2475 / ajs.s3-37.220.323. S2CID 130946518.
- Holland, W. J. (1919). «V бөлім. Палеонтология». 1919 жылдың 31 наурызында аяқталатын жылға арналған директордың жылдық есебі (Есеп). Карнеги мұражайы. б. 38. hdl:2027 / osu.32435027945948.
- Holland, W. J. (1924). «Uintasaurus douglassi HOLLAND типінің сипаттамасы». Карнеги мұражайының жылнамалары. 15 (2–3): 119–138.
- Осборн, Х. Ф .; Mook, C. C. (1921). «Камарасавр, Амфикоэлия және Коптың басқа да сауоподтары». Американың геологиялық қоғамының хабаршысы. 30 (1): 379–388. Бибкод:1919GSAB ... 30..379O. дои:10.1130 / GSAB-30-379. hdl:2027 / mdp.39015042532476.
- Элингер, Т. Х. (1950). «Camarosaurus annae - жаңа американдық сауопод динозавр». Американдық натуралист. 84 (816): 225–228. дои:10.1086/281626.
- Дженсен, Дж. А. (1988). «Колорадо үстіртінің жоғарғы юра дәуірінен шыққан төртінші жаңа сауопод динозавры және сауропод бипедализмі». Ұлы бассейн натуралисті. 48 (2): 121–145. JSTOR 41712420.
- Макинтош, Дж. С .; Миллер, В. Штадман, К.Л .; Джиллетт, Д.Д (1996). «Camarasaurus lewisi остеологиясы (Дженсен, 1988)». Бригам Янг университетінің геологиялық зерттеулер. 41: 73–95.
- Макинтош, Дж. С .; Майлз, С .; Cloward, K. C .; Паркер, Дж. Р. (1996). «Камарасаврдың толықтай жаңа қаңқасы». Гунма табиғи мұражайының хабаршысы (1): 1–87.
- Додсон, Питер; Бритт, Брукс; Ағаш ұстасы, Кеннет; Форстер, Кэтрин А .; Джилетт, Дэвид Д .; Норелл, Марк А .; Ольшевский, Джордж; Париш, Дж. Майкл; Вейшампел, Дэвид Б. (1993-01-01). Динозаврлар дәуірі. Publications International, LTD. ISBN 978-0-7853-0443-2.
- Фостер, Дж. (2007). Юра Батысы: Моррисон формациясының динозаврлары және олардың әлемі. Индиана университетінің баспасы. ISBN 978-0-253-34870-8.
- Тидуэлл, V .; Ағаш ұстасы, К .; Meyer, S. (2001). Танке, Д. Х .; Ағаш ұстасы, К. (ред.). «Жаңа Ситан тауриформасы (Сауропода), Сидар тау түзілісінің (Төменгі бор), Юта штаты мүшесінен». Мезозой омыртқалы тіршілігі: 139–165.
- Қанаттар, Оливер; Sander, P. Martin (2007). «Сауропод динозаврларында асқазан диірмені жоқ: гастролит массасы мен түйеқұстардағы функциялардың анализі» (PDF). Корольдік қоғамның еңбектері B. 274 (1610): 635–640. дои:10.1098 / rspb.2006.3763. PMC 2197205. PMID 17254987. Архивтелген түпнұсқа (PDF) 2011-07-13. Алынған 2009-07-24.
- Маквинни, Лорри; Ағаш ұстасы, Кеннет; Ротшильд, Брюс (2001), «25. Динозаврлық гумеральды периостит: қазба жазбаларындағы юкстакортикальды зақымдану жағдайы», Мезозой омыртқалы тіршілігі, Индиана университетінің баспасы, 364–377 бб
Сыртқы сілтемелер
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||












